

Recruitment of the NineTeen Complex to the activated spliceosome requires AtPRMT5
- PMID: 27114555
- PMCID: PMC4868449
- DOI: 10.1073/pnas.1522458113
Recruitment of the NineTeen Complex to the activated spliceosome requires AtPRMT5
Erratum in
-
Correction to Supporting Information for Deng et al., Recruitment of the NineTeen Complex to the activated spliceosome requires AtPRMT5.Proc Natl Acad Sci U S A. 2016 May 31;113(22):E3186. doi: 10.1073/pnas.1607263113. Epub 2016 May 23. Proc Natl Acad Sci U S A. 2016. PMID: 27217555 Free PMC article. No abstract available.
Abstract
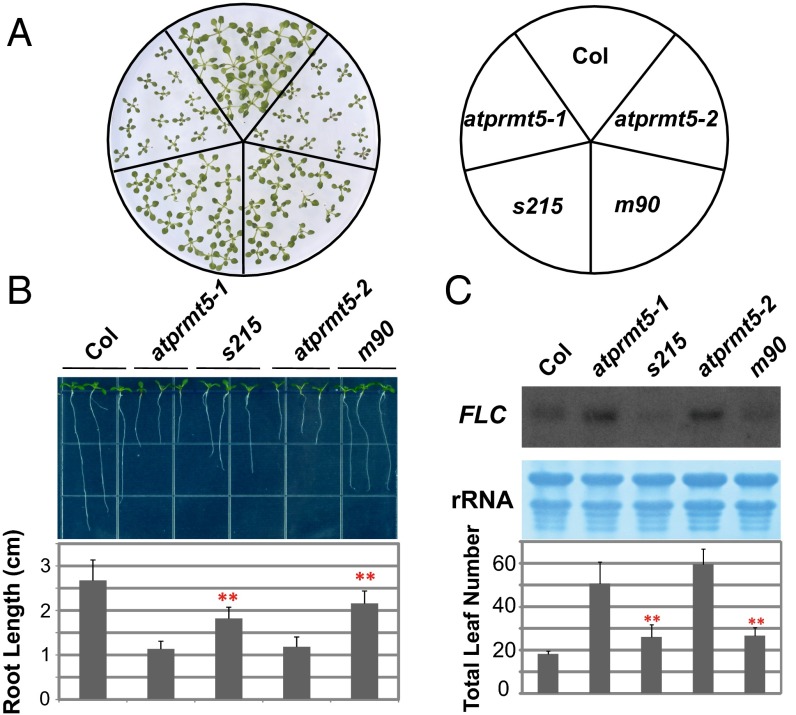
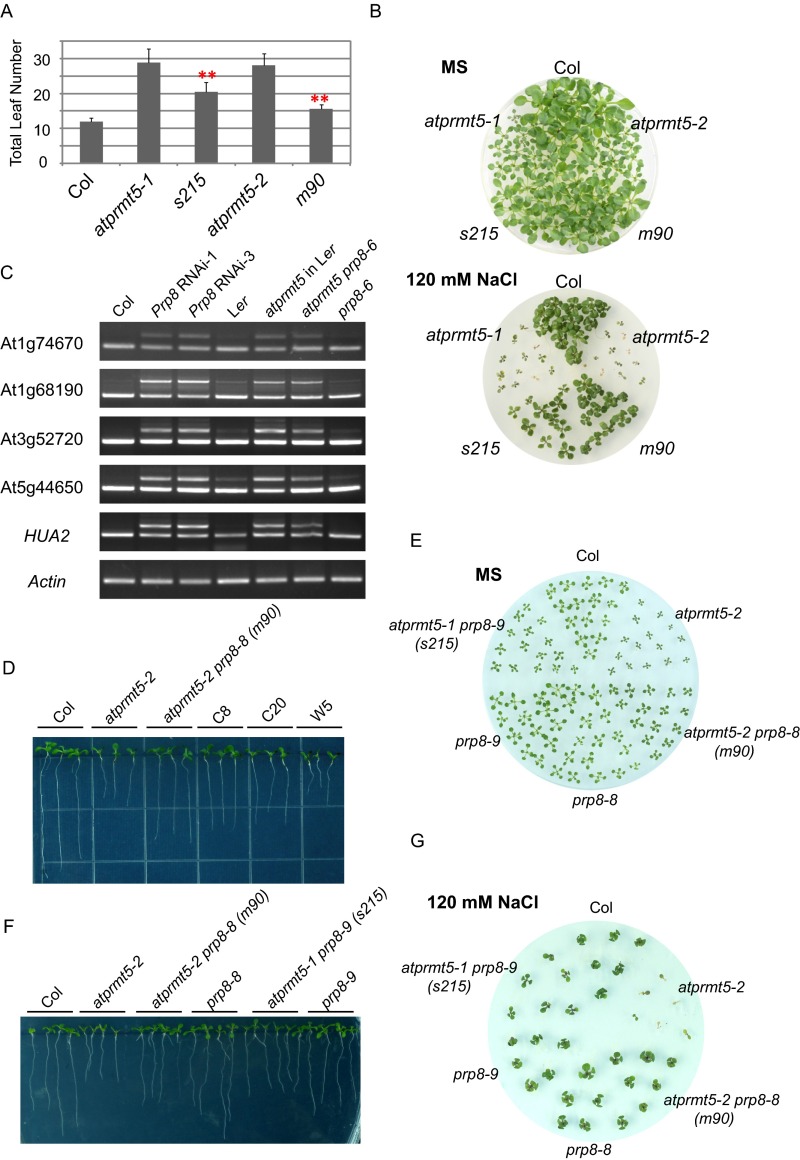
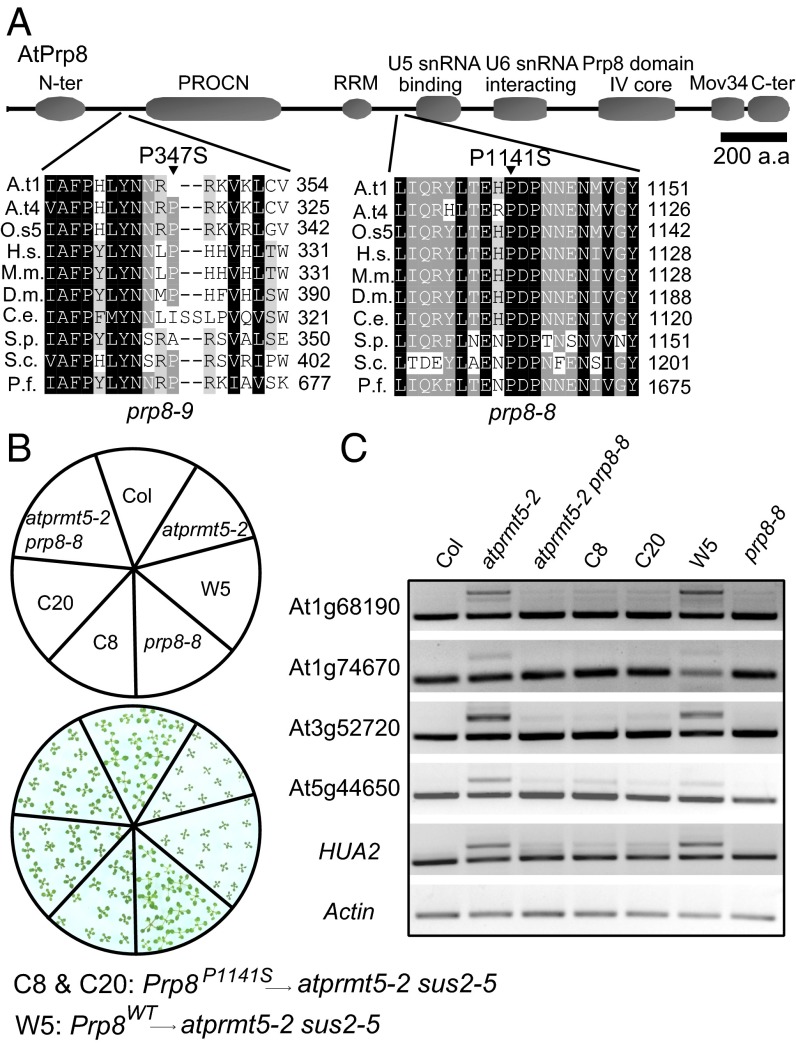
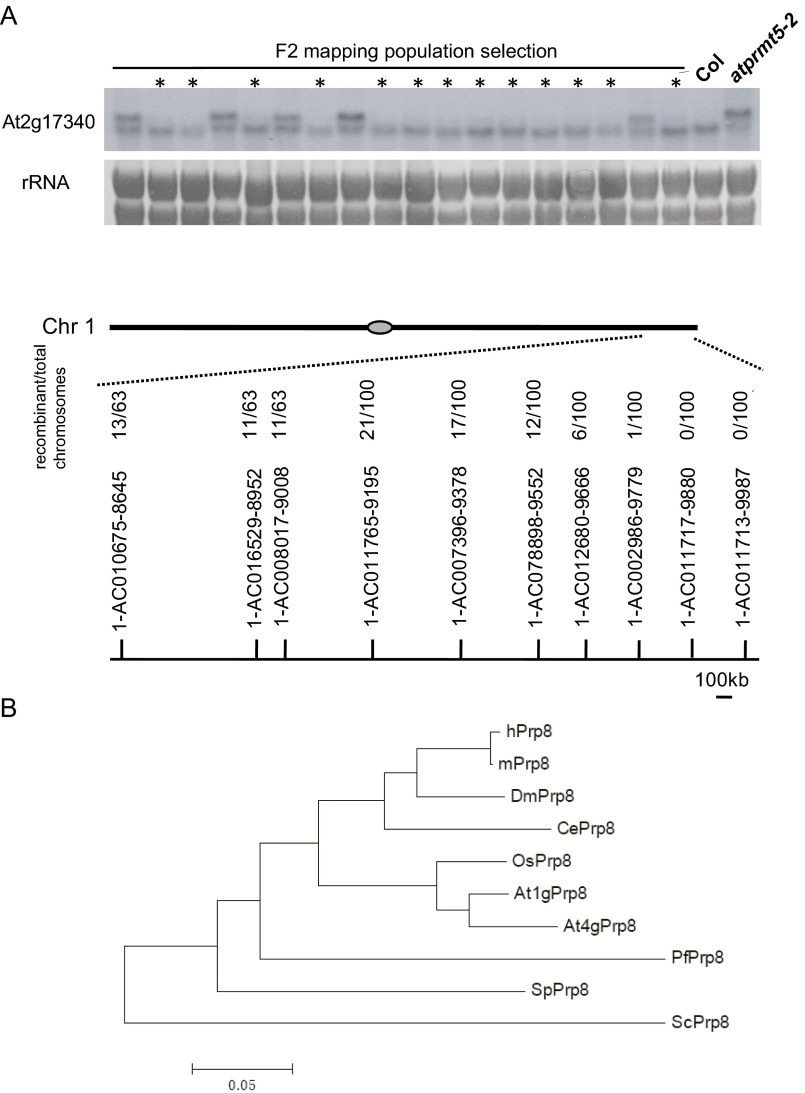
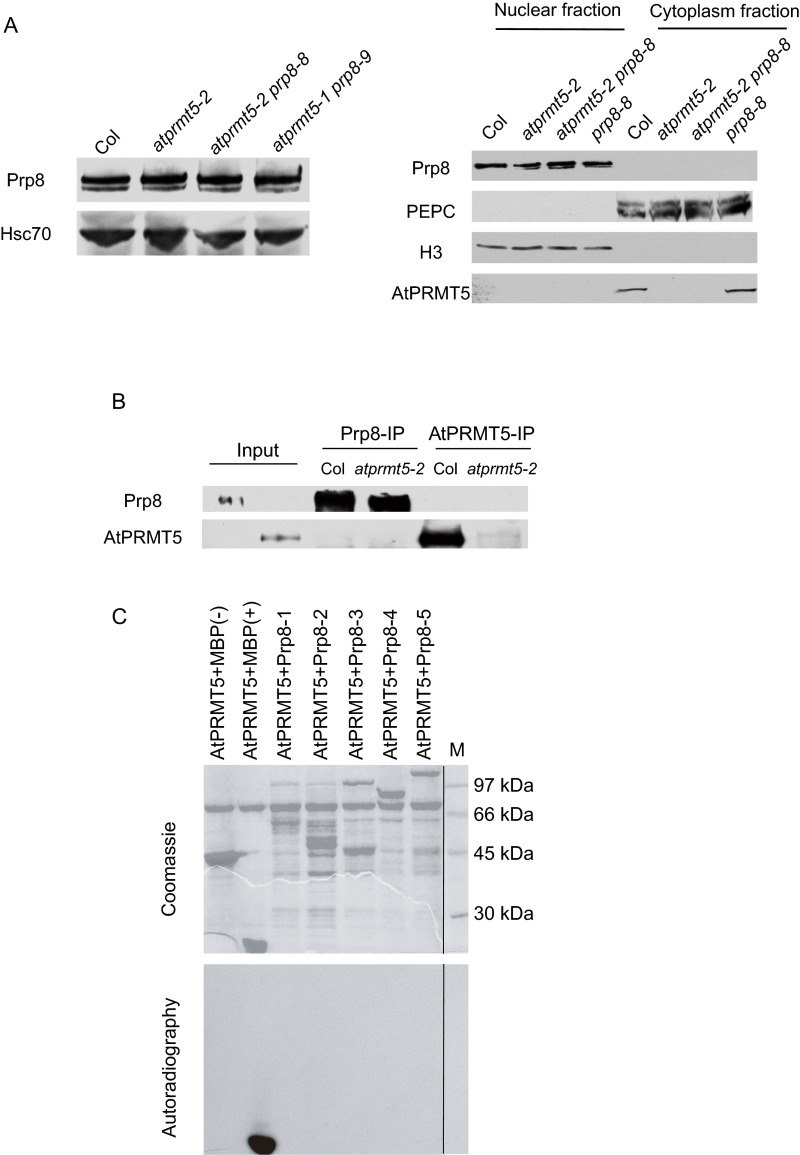
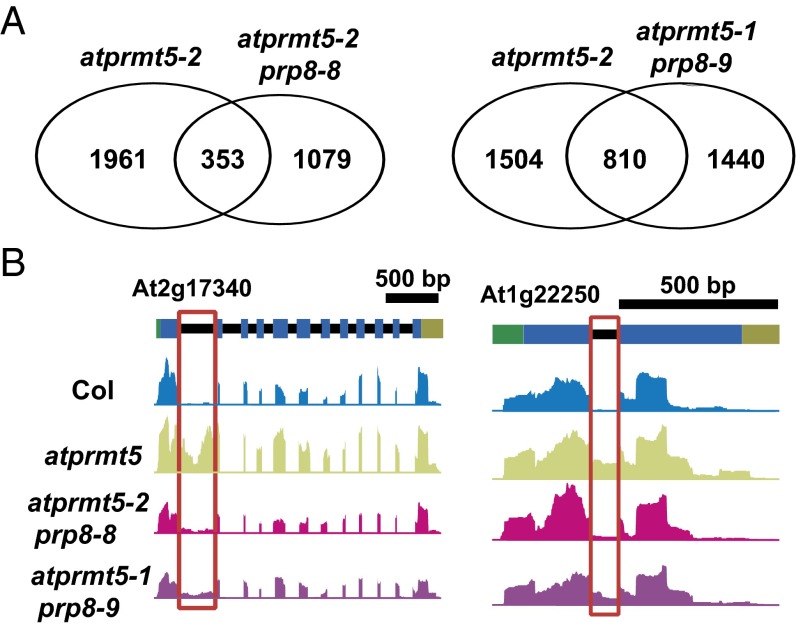
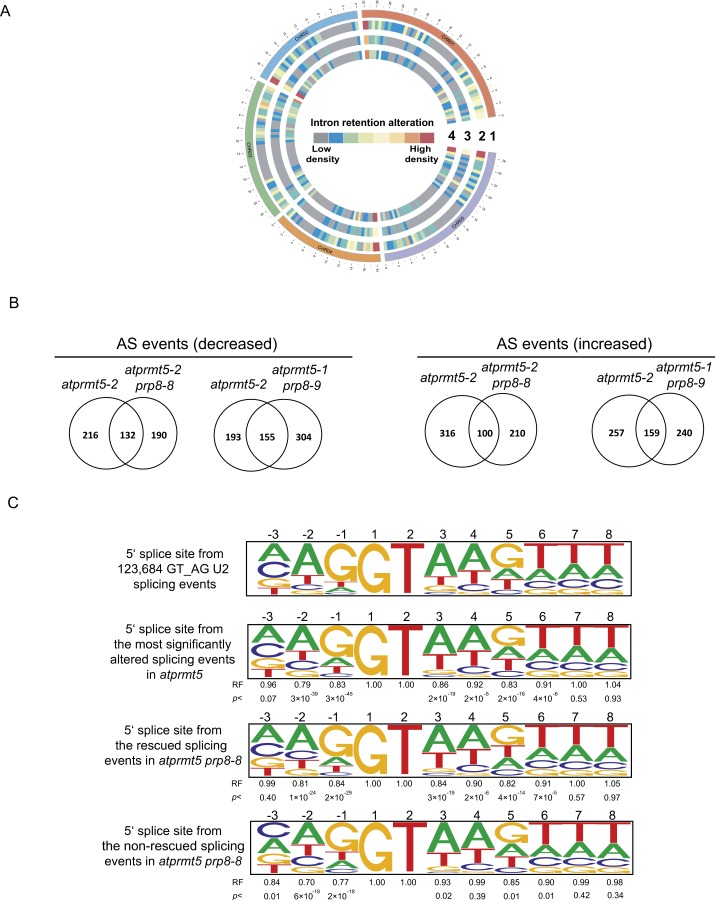
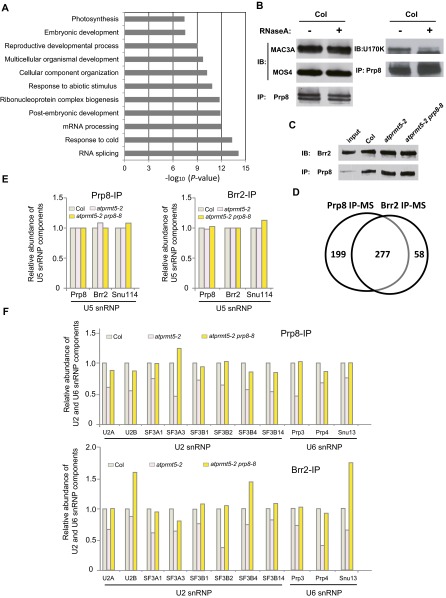
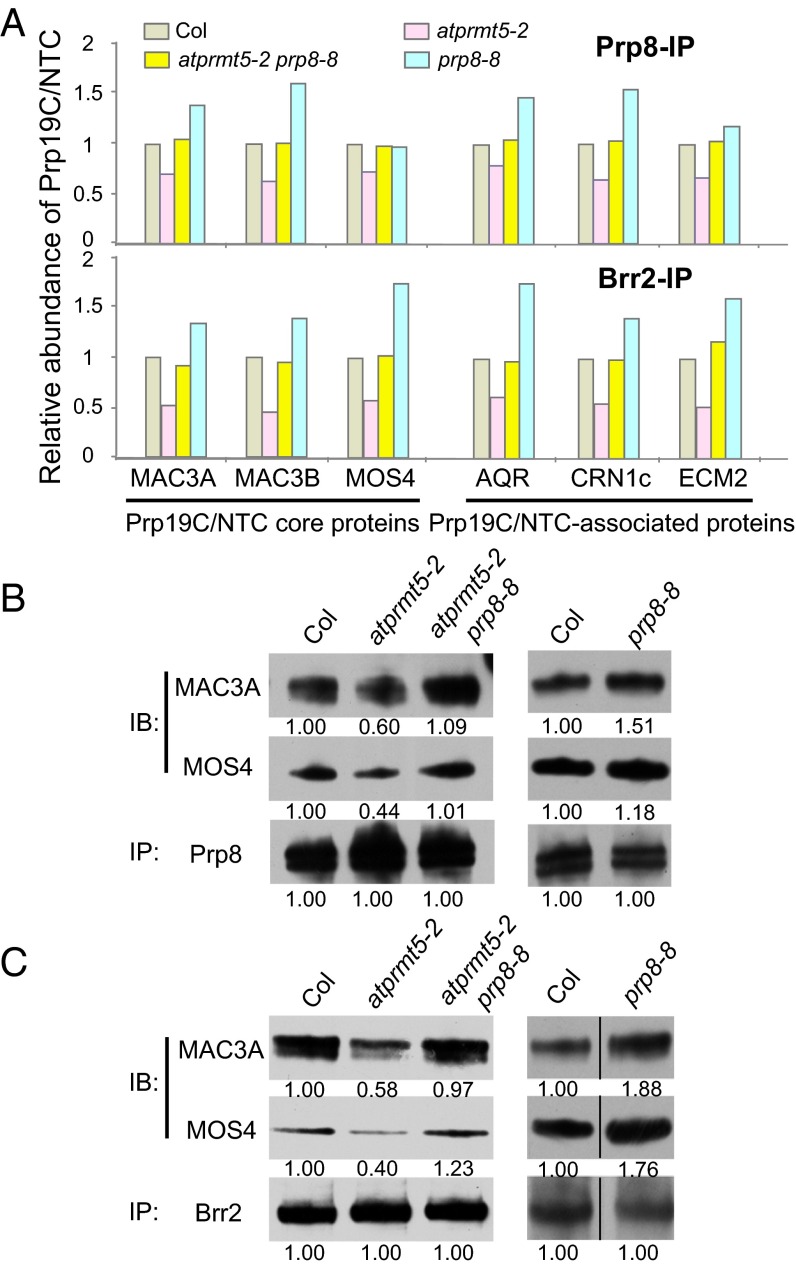
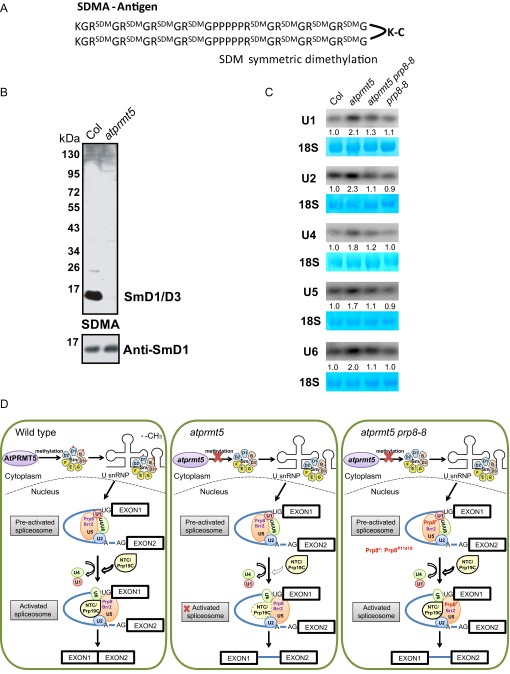
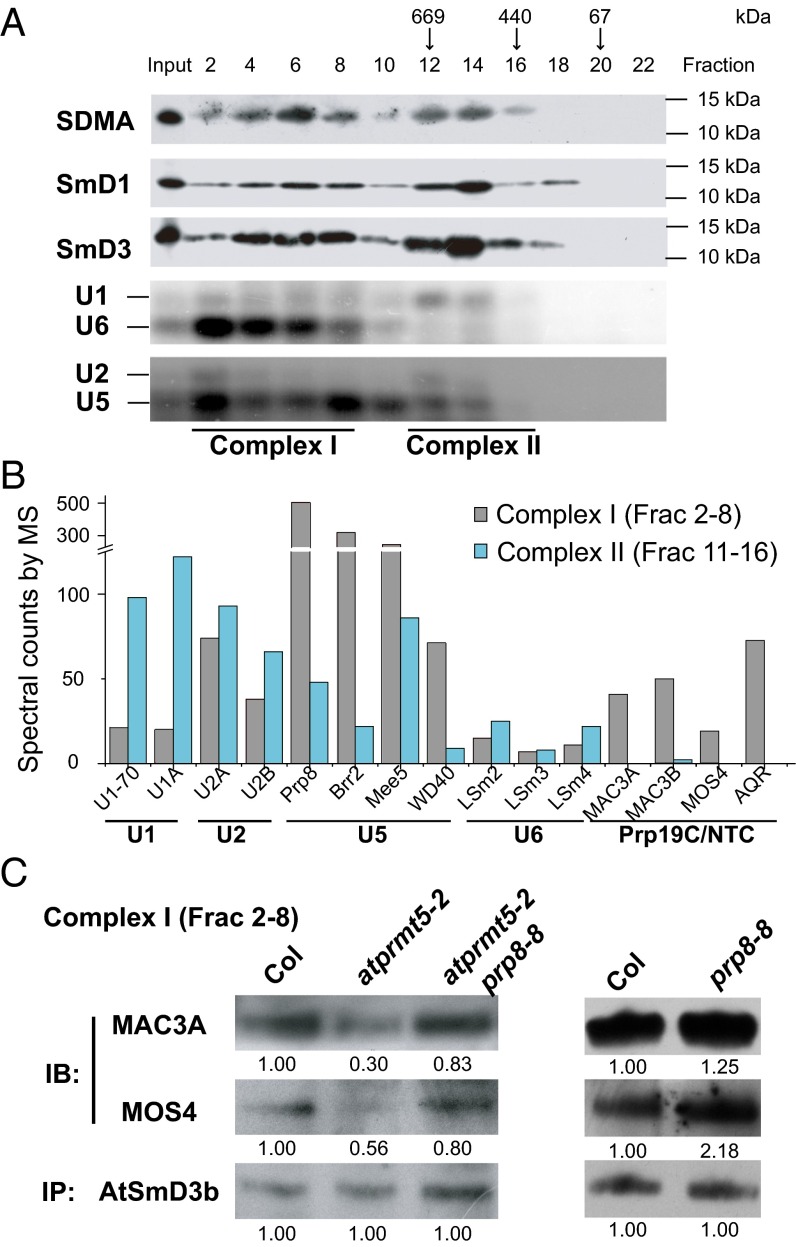
Protein arginine methylation, catalyzed by protein arginine methyltransferases (PRMTs), is involved in a multitude of biological processes in eukaryotes. Symmetric arginine dimethylation mediated by PRMT5 modulates constitutive and alternative pre-mRNA splicing of diverse genes to regulate normal growth and development in multiple species; however, the underlying molecular mechanism remains largely unknown. A genetic screen for suppressors of an Arabidopsis symmetric arginine dimethyltransferase mutant, atprmt5, identified two gain-of-function alleles of pre-mRNA processing factor 8 gene (prp8-8 and prp8-9), the highly conserved core component of the U5 small nuclear ribonucleoprotein (snRNP) and the spliceosome. These two atprmt5 prp8 double mutants showed suppression of the developmental and splicing alterations of atprmt5 mutants. In atprmt5 mutants, the NineTeen complex failed to be assembled into the U5 snRNP to form an activated spliceosome; this phenotype was restored in the atprmt5 prp8-8 double mutants. We also found that loss of symmetric arginine dimethylation of Sm proteins prevents recruitment of the NineTeen complex and initiation of spliceosome activation. Together, our findings demonstrate that symmetric arginine dimethylation has important functions in spliceosome assembly and activation, and uncover a key molecular mechanism for arginine methylation in pre-mRNA splicing that impacts diverse developmental processes.
Keywords: AtPRMT5; Prp19C/NTC; arginine methylation; pre-mRNA splicing; protein arginine methyltransferase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Arginine methylation mediated by the Arabidopsis homolog of PRMT5 is essential for proper pre-mRNA splicing.Proc Natl Acad Sci U S A. 2010 Nov 2;107(44):19114-9. doi: 10.1073/pnas.1009669107. Epub 2010 Oct 18. Proc Natl Acad Sci U S A. 2010. PMID: 20956294 Free PMC article.
-
AtPRMT5 Regulates Shoot Regeneration through Mediating Histone H4R3 Dimethylation on KRPs and Pre-mRNA Splicing of RKP in Arabidopsis.Mol Plant. 2016 Dec 5;9(12):1634-1646. doi: 10.1016/j.molp.2016.10.010. Epub 2016 Oct 22. Mol Plant. 2016. PMID: 27780782
-
The splicing factor Prp17 interacts with the U2, U5 and U6 snRNPs and associates with the spliceosome pre- and post-catalysis.Biochem J. 2008 Dec 15;416(3):365-74. doi: 10.1042/BJ20081195. Biochem J. 2008. PMID: 18691155
-
Functional roles of DExD/H-box RNA helicases in Pre-mRNA splicing.J Biomed Sci. 2015 Jul 16;22(1):54. doi: 10.1186/s12929-015-0161-z. J Biomed Sci. 2015. PMID: 26173448 Free PMC article. Review.
-
Splicing and beyond: the many faces of the Prp19 complex.Biochim Biophys Acta. 2013 Oct;1833(10):2126-34. doi: 10.1016/j.bbamcr.2013.05.023. Epub 2013 Jun 3. Biochim Biophys Acta. 2013. PMID: 23742842 Review.
Cited by
-
Light controls mesophyll-specific post-transcriptional splicing of photoregulatory genes by AtPRMT5.Proc Natl Acad Sci U S A. 2024 Feb 6;121(6):e2317408121. doi: 10.1073/pnas.2317408121. Epub 2024 Jan 29. Proc Natl Acad Sci U S A. 2024. PMID: 38285953 Free PMC article.
-
Alternative splicing as a key player in the fine-tuning of the immunity response in Arabidopsis.Mol Plant Pathol. 2022 Aug;23(8):1226-1238. doi: 10.1111/mpp.13228. Epub 2022 May 14. Mol Plant Pathol. 2022. PMID: 35567423 Free PMC article. Review.
-
Exogenous Auxin Elicits Changes in the Arabidopsis thaliana Root Proteome in a Time-Dependent Manner.Proteomes. 2017 Jul 10;5(3):16. doi: 10.3390/proteomes5030016. Proteomes. 2017. PMID: 28698516 Free PMC article.
-
Recent advances in the regulation of plant miRNA biogenesis.RNA Biol. 2021 Dec;18(12):2087-2096. doi: 10.1080/15476286.2021.1899491. Epub 2021 Mar 17. RNA Biol. 2021. PMID: 33666136 Free PMC article. Review.
-
XAP5 CIRCADIAN TIMEKEEPER regulates RNA splicing and the circadian clock by genetically separable pathways.Plant Physiol. 2023 Jul 3;192(3):2492-2506. doi: 10.1093/plphys/kiad193. Plant Physiol. 2023. PMID: 36974904 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
