

Dysregulation of miRNA-9 in a Subset of Schizophrenia Patient-Derived Neural Progenitor Cells
- PMID: 27117414
- PMCID: PMC4856588
- DOI: 10.1016/j.celrep.2016.03.090
Dysregulation of miRNA-9 in a Subset of Schizophrenia Patient-Derived Neural Progenitor Cells
Erratum in
-
Dysregulation of miRNA-9 in a Subset of Schizophrenia Patient-Derived Neural Progenitor Cells.Cell Rep. 2017 Sep 5;20(10):2525. doi: 10.1016/j.celrep.2017.08.073. Cell Rep. 2017. PMID: 28877483 Free PMC article. No abstract available.
Abstract
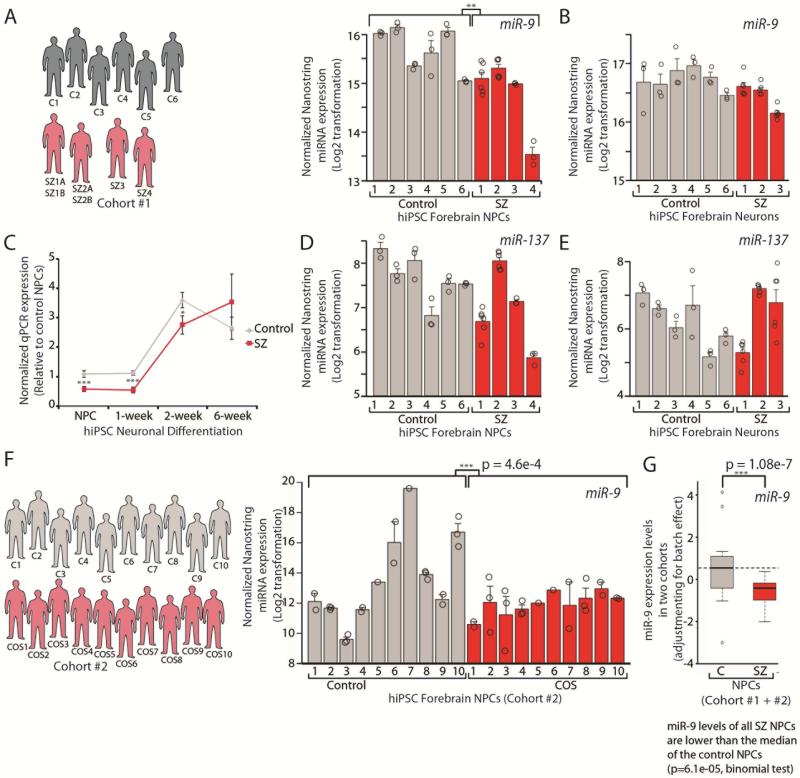
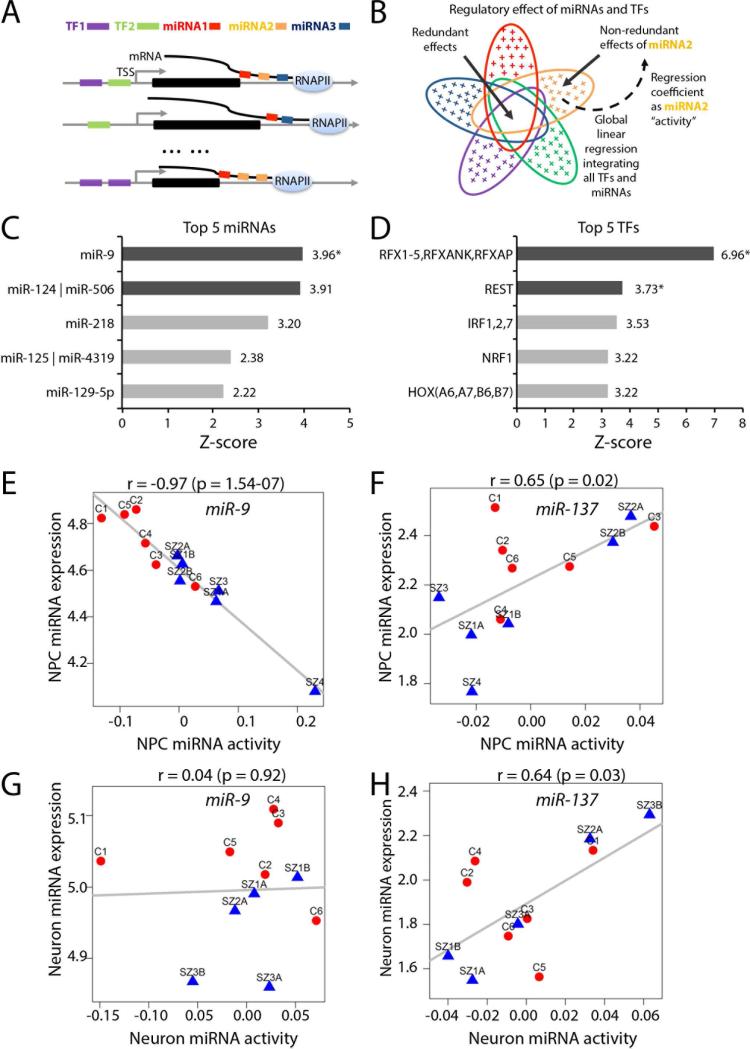
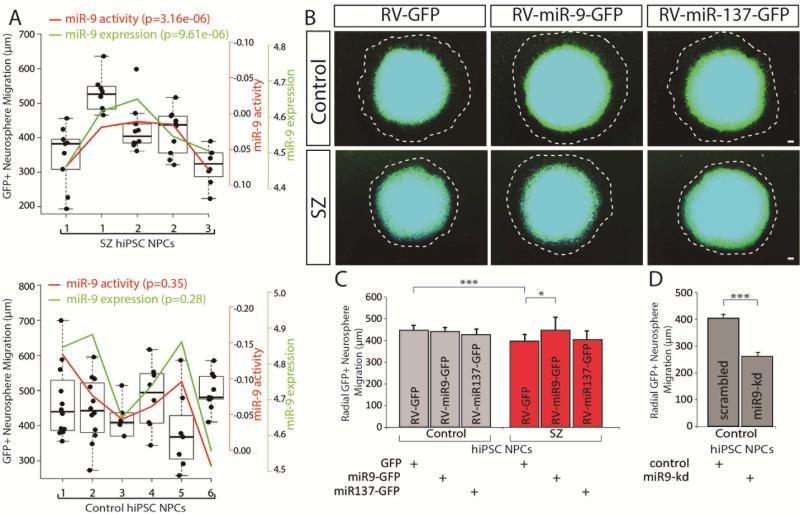
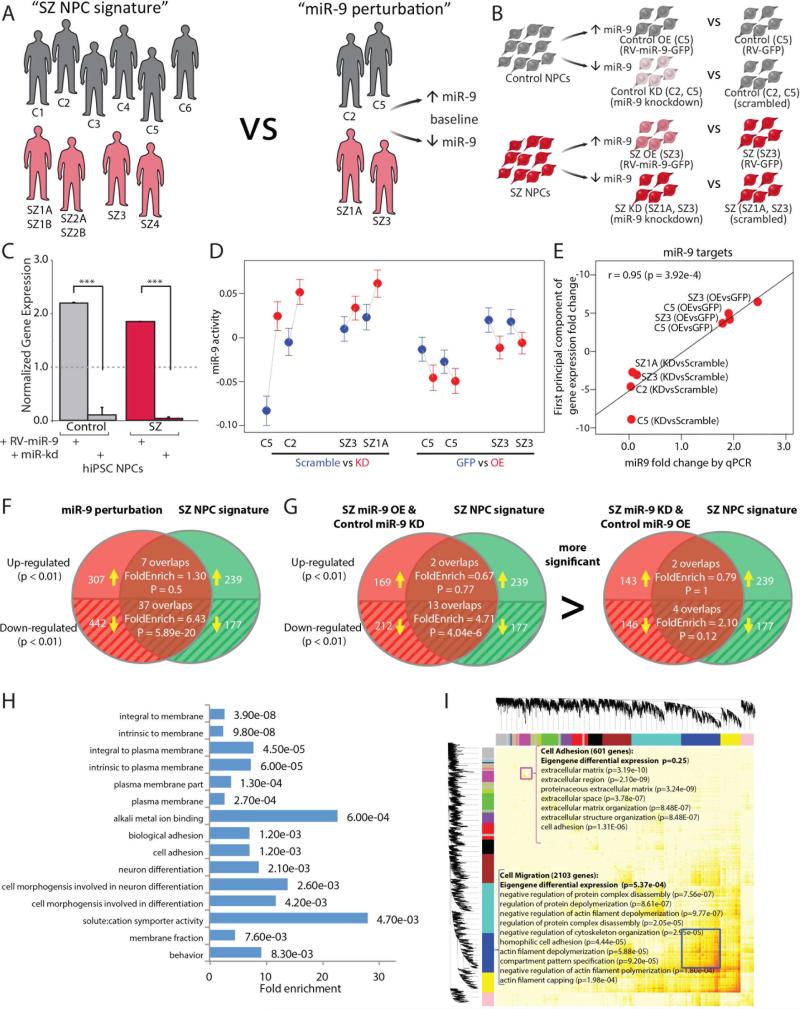
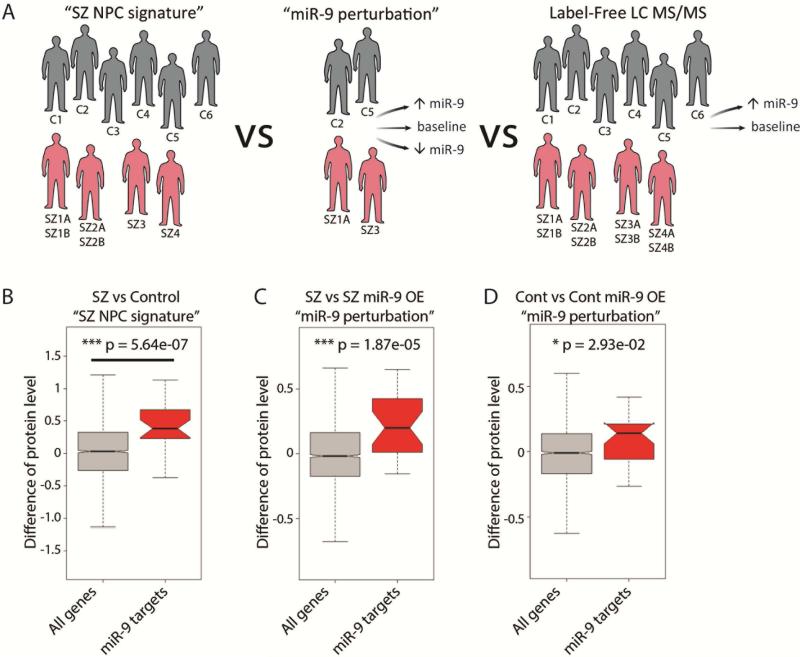
Converging evidence indicates that microRNAs (miRNAs) may contribute to disease risk for schizophrenia (SZ). We show that microRNA-9 (miR-9) is abundantly expressed in control neural progenitor cells (NPCs) but also significantly downregulated in a subset of SZ NPCs. We observed a strong correlation between miR-9 expression and miR-9 regulatory activity in NPCs as well as between miR-9 levels/activity, neural migration, and diagnosis. Overexpression of miR-9 was sufficient to ameliorate a previously reported neural migration deficit in SZ NPCs, whereas knockdown partially phenocopied aberrant migration in control NPCs. Unexpectedly, proteomic- and RNA sequencing (RNA-seq)-based analysis revealed that these effects were mediated primarily by small changes in expression of indirect miR-9 targets rather than large changes in direct miR-9 targets; these indirect targets are enriched for migration-associated genes. Together, these data indicate that aberrant levels and activity of miR-9 may be one of the many factors that contribute to SZ risk, at least in a subset of patients.
Keywords: human-induced pluripotent stem cell; microRNA-9; neural progenitor cells; schizophrenia.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
