

IL-27 Promotes Proliferation of Human Leukemic Cell Lines Through the MAPK/ERK Signaling Pathway and Suppresses Sensitivity to Chemotherapeutic Drugs
- PMID: 27119567
- PMCID: PMC4854210
- DOI: 10.1089/jir.2015.0091
IL-27 Promotes Proliferation of Human Leukemic Cell Lines Through the MAPK/ERK Signaling Pathway and Suppresses Sensitivity to Chemotherapeutic Drugs
Abstract
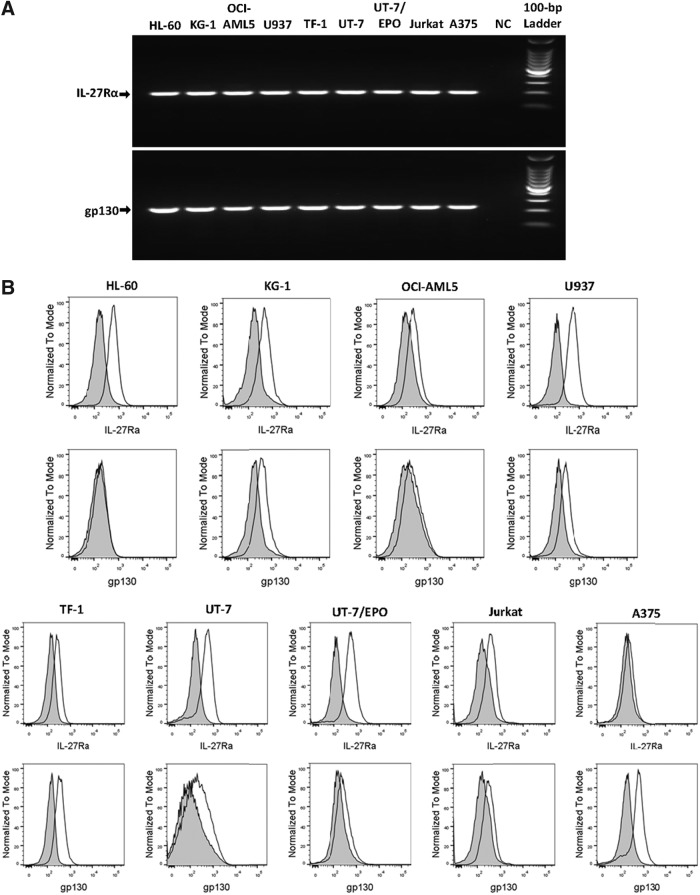
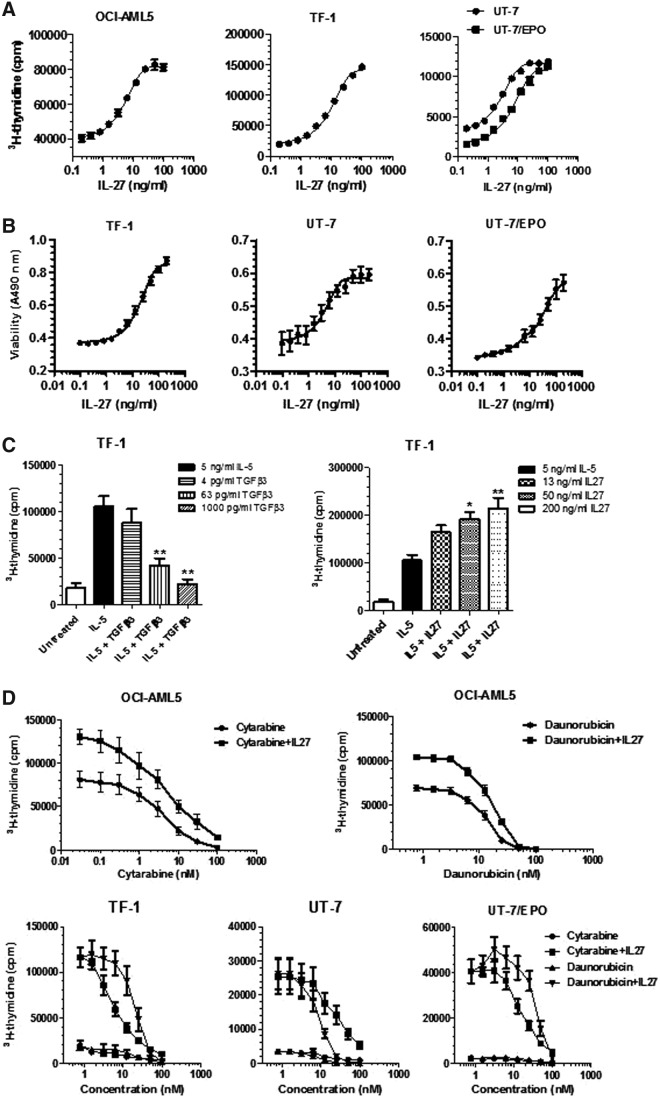
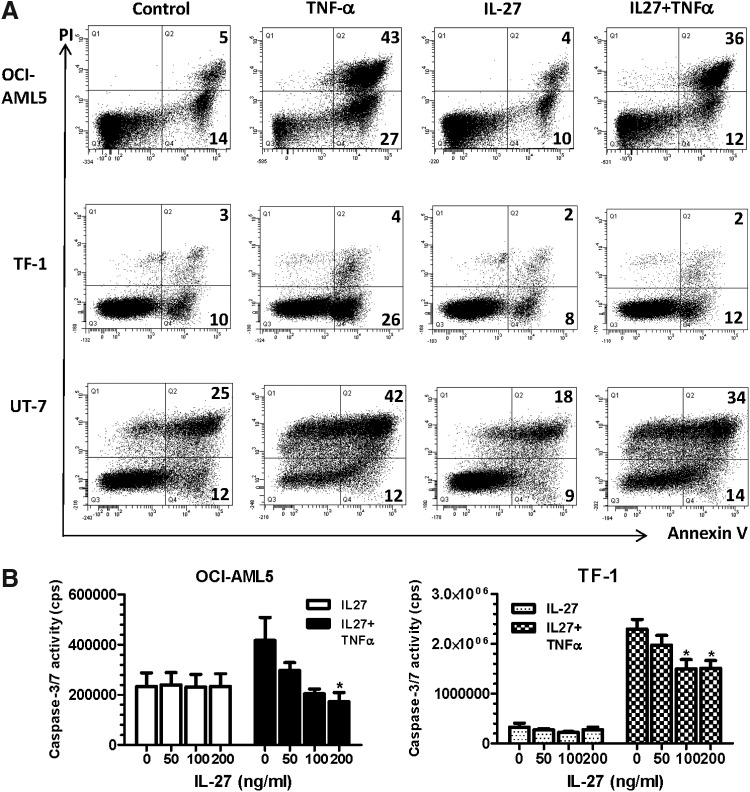
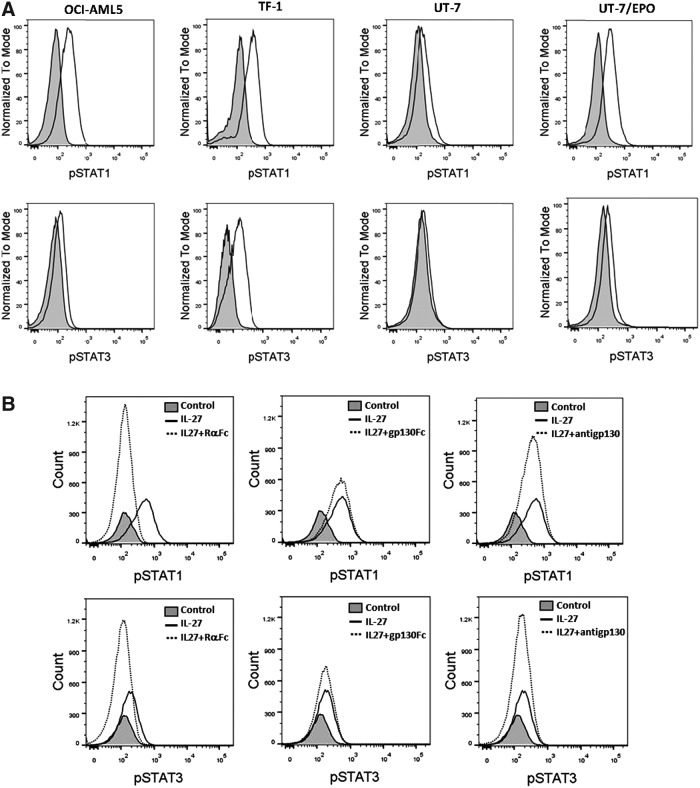
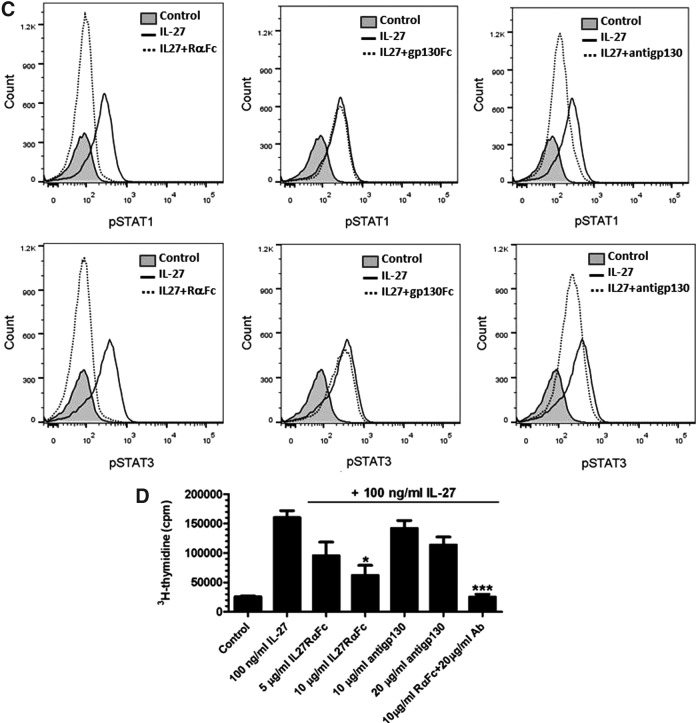
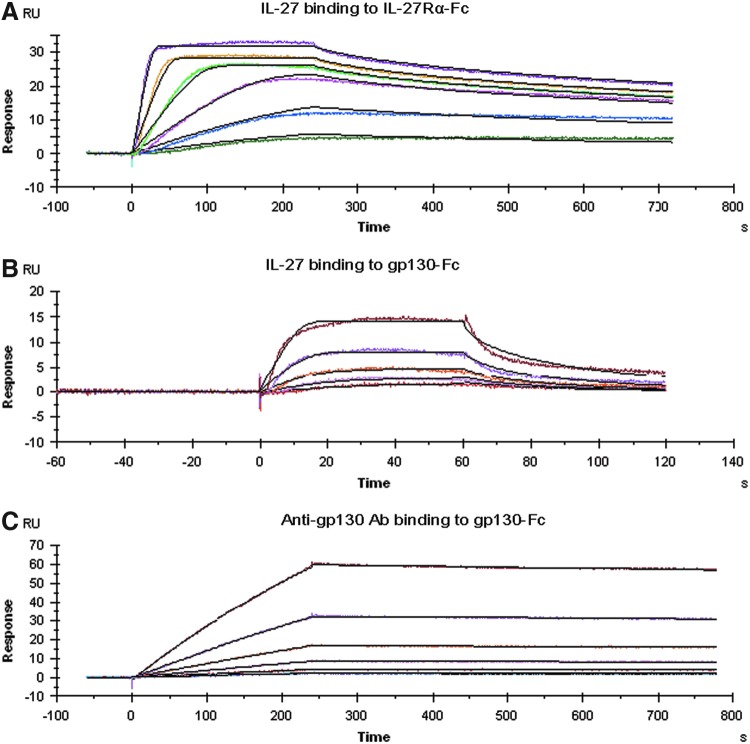
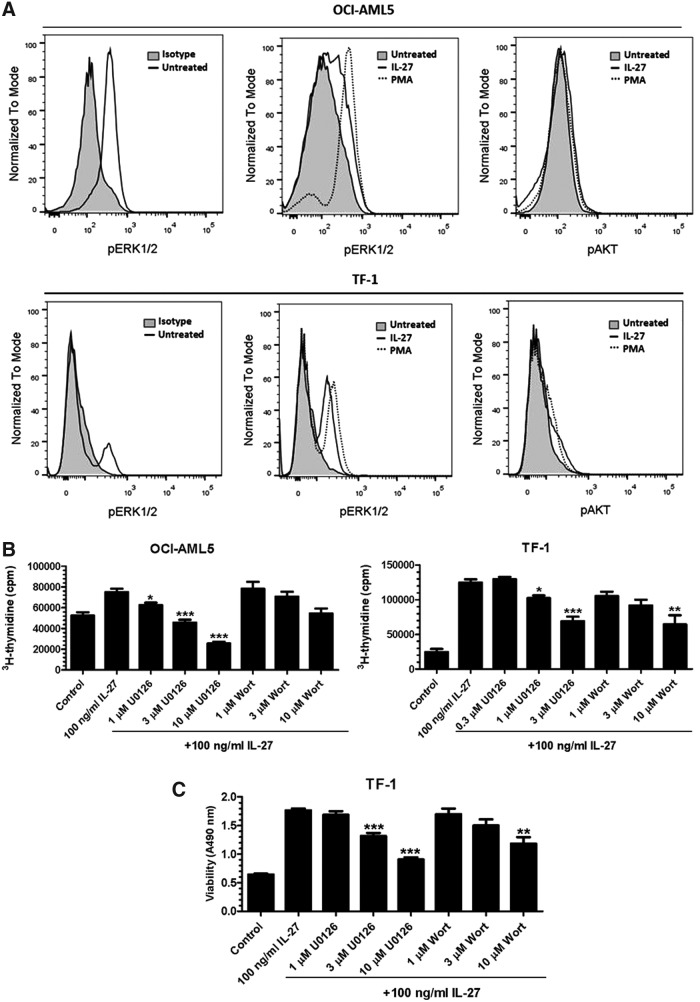
IL-27 is a pleiotropic cytokine of the IL-6/IL-12 family with diverse biological functions. Previous in vivo studies have suggested the antitumor activities of IL-27 in animal models, whereas clinical observations indicate the link of IL-27 in tumor progression. IL-27 has recently been shown to cause inhibition of proliferation on primary leukemic cells from pediatric patients, but information on its role in human leukemic cell lines is limited. In the present study, we investigated the ability of IL-27 to regulate cell growth and survival of various human leukemic cell lines. Our results showed that in human leukemic cell lines coexpressing both IL-27R chains, IL-27Rα and gp130, IL-27 did not inhibit cell growth, but caused dose-dependent proliferation of the acute myeloid leukemic cell line, OCI-AML5, and the erythroleukemic cell lines, TF-1, UT-7, and UT-7/EPO. Consistent with this, IL-27 promoted cell survival and reduced TNF-α-induced apoptosis of the leukemic cell lines. IL-27 also decreased the responsiveness of the leukemic cells to chemotherapeutic drugs, cytarabine and daunorubicin. We observed that IL-27 induced the activation of STAT1/3 and ERK1/2 in the leukemic cells. Growth stimulation by IL-27 was suppressed by the specific MEK inhibitor, U0126, indicating that IL-27-induced cell proliferation is mainly mediated through the activation of the MAPK/ERK signaling pathway. The present study is the first demonstration of the proliferative and antichemotherapeutic properties of IL-27 in human leukemic cell lines, suggesting that IL-27 can play an unfavorable role in tumor growth and can be an important determinant in the chemoresponsiveness of certain subtypes of human leukemia.
Figures
References
-
- Awasthi A, Carrier Y, Peron JP, Bettelli E, Kamanaka M, Flavell RA, Kuchroo VK, Oukka M, Weiner HL. 2007. A dominant function for interleukin 27 in generating interleukin 10-producing anti-inflammatory T cells. Nat Immunol 8:1380–1389 - PubMed
-
- Canale S, Cocco C, Frasson C, Seganfreddo E, Di Carlo E, Ognio E, Sorrentino C, Ribatti D, Zorzoli A, Basso G, Dufour C, Airoldi I. 2011. Interleukin-27 inhibits pediatric B-acute lymphoblastic leukemia cell spreading in a preclinical model. Leukemia 25:1815–1824 - PubMed
-
- Creutzig U, van den Heuvel-Eibrink MM, Gibson B, Dworzak MN, Adachi S, de Bont E, Harbott J, Hasle H, Johnston D, Kinoshita A, Lehrnbecher T, Leverger G, Mejstrikova E, Meshinchi S, Pession A, Raimondi SC, Sung L, Stary J, Zwaan CM, Kaspers GJ, Reinhardt D. 2012. AML Committee of the International BFM Study Group. Diagnosis and management of acute myeloid leukemia in children and adolescents: recommendations from an international expert panel. Blood 120:3187–3205 - PubMed
-
- Diakowska D, Lewandowski A, Markocka-Mączka K, GrAbowski K. 2013. Concentration of serum interleukin-27 increase in patients with lymph node metastatic gastroesophageal cancer. Adv Clin Exp Med 22:683–691 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous