

Comparative Phylogeography Reveals Cryptic Diversity and Repeated Patterns of Cladogenesis for Amphibians and Reptiles in Northwestern Ecuador
- PMID: 27120100
- PMCID: PMC4847877
- DOI: 10.1371/journal.pone.0151746
Comparative Phylogeography Reveals Cryptic Diversity and Repeated Patterns of Cladogenesis for Amphibians and Reptiles in Northwestern Ecuador
Abstract
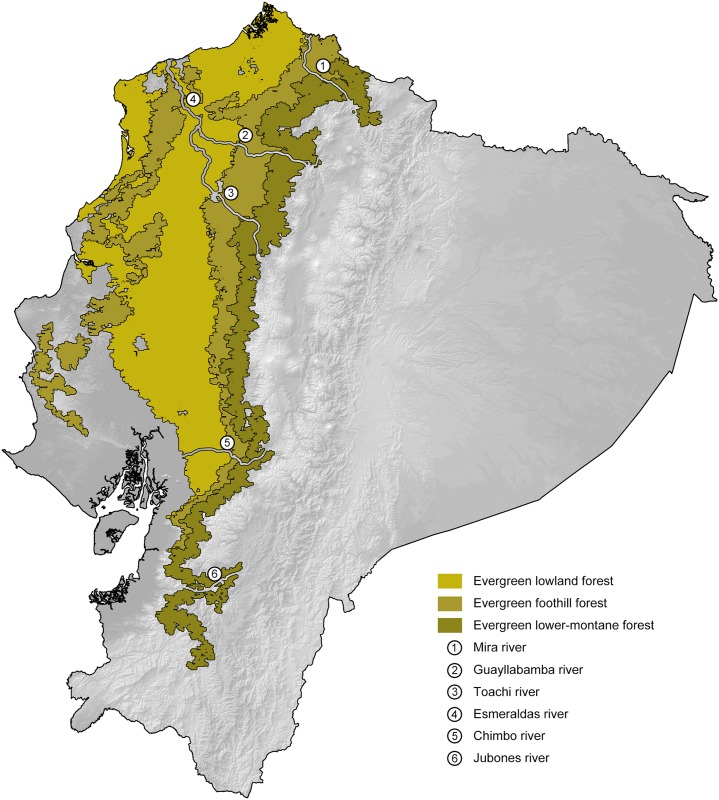
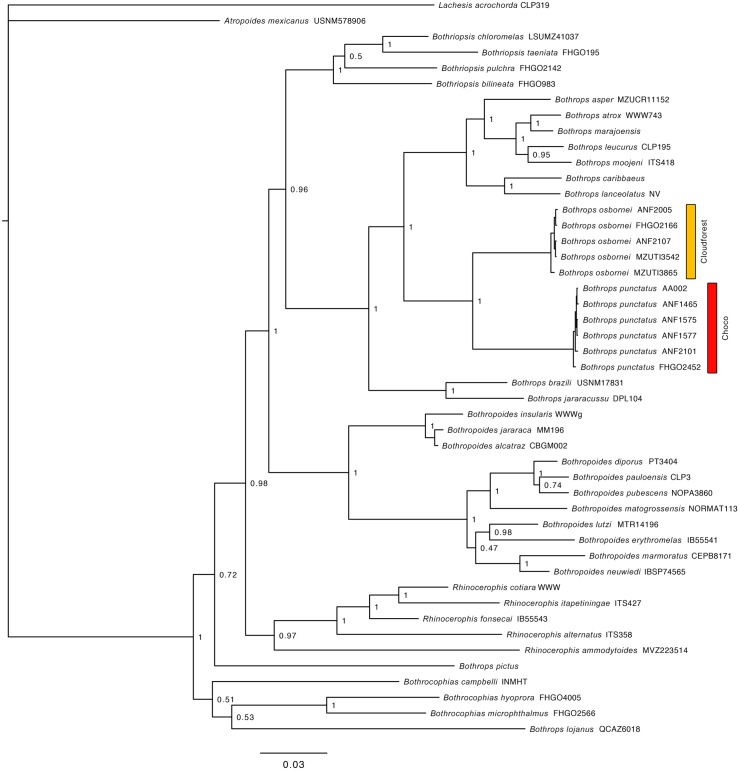
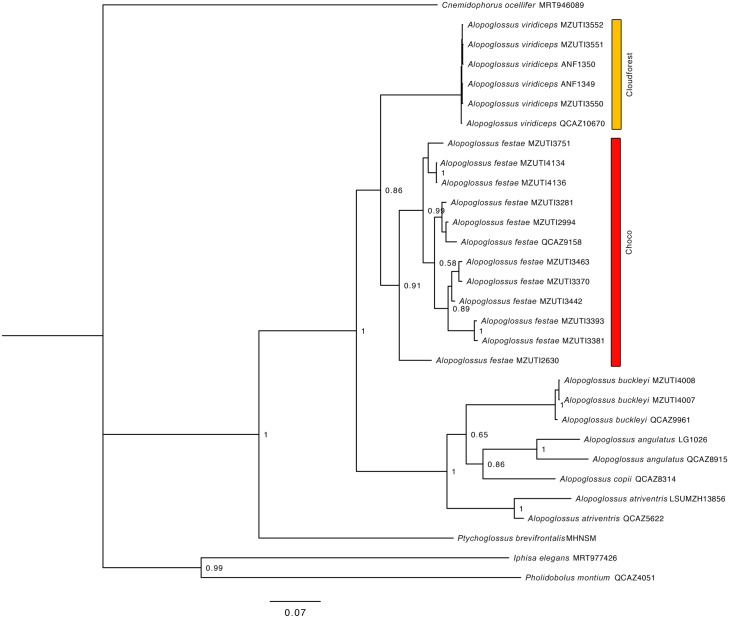
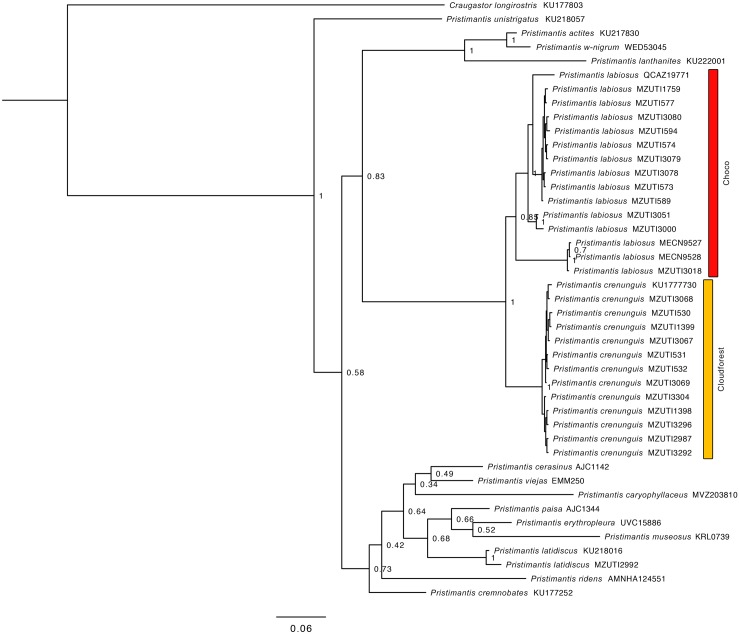
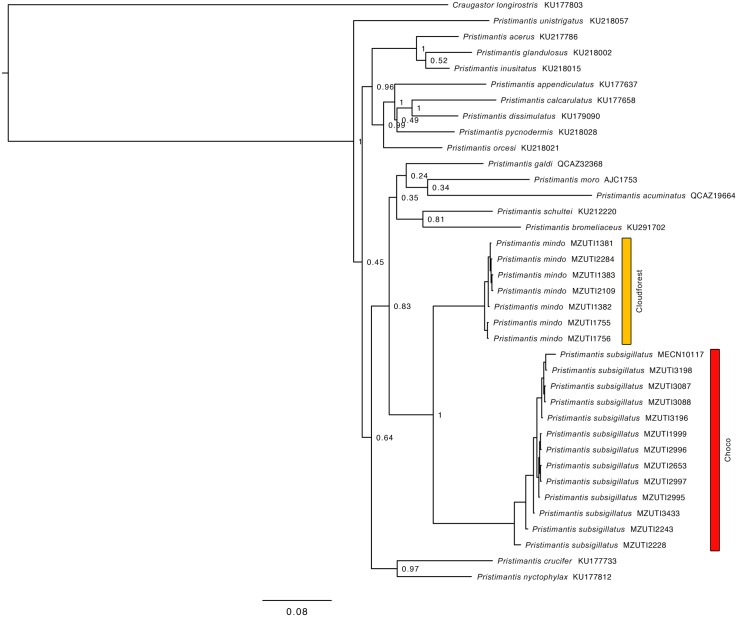
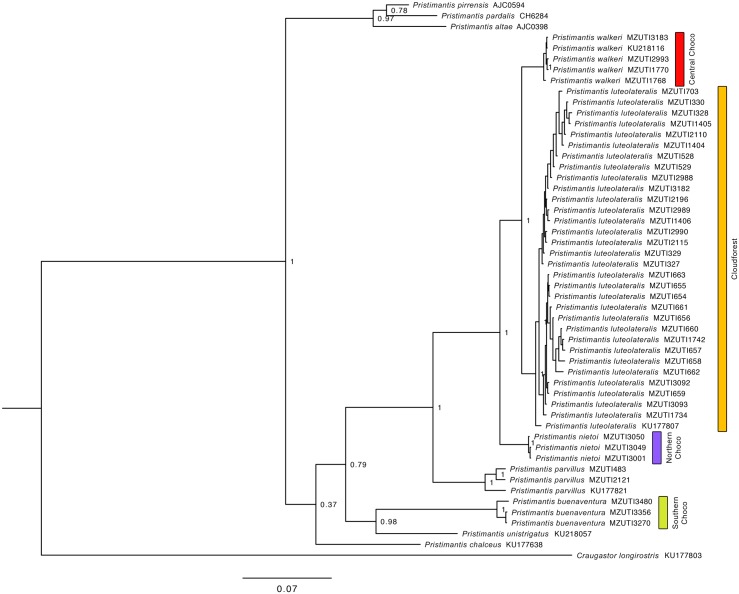
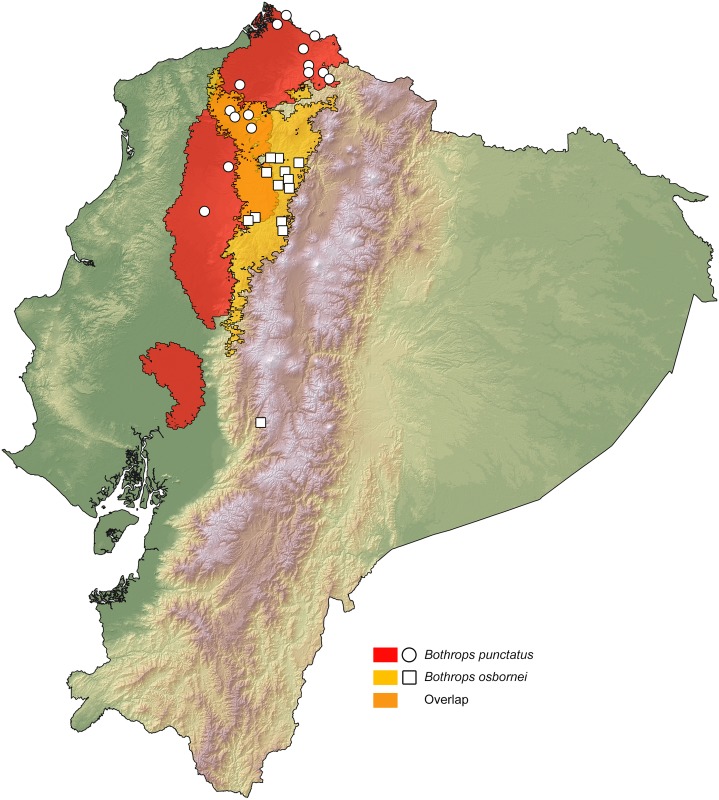
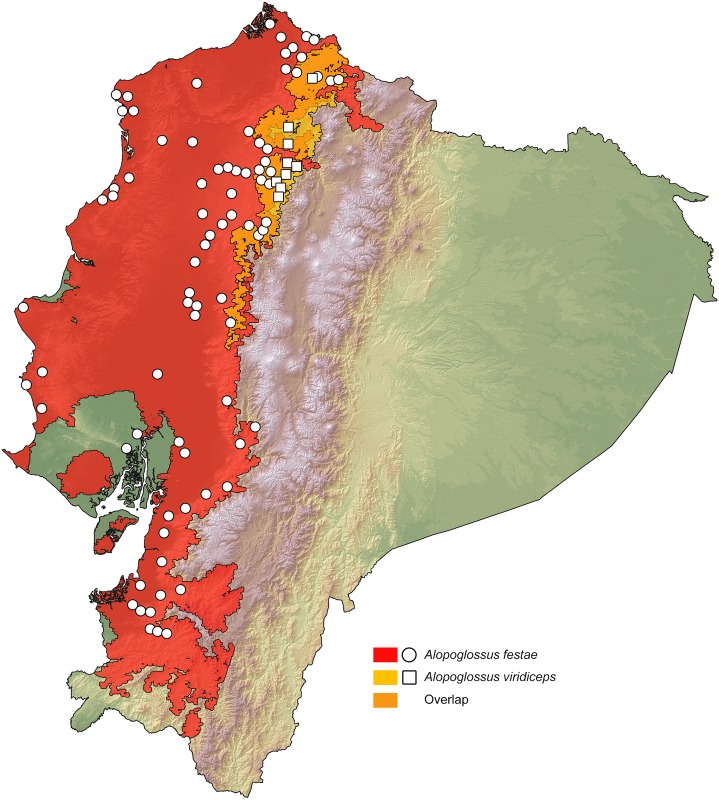
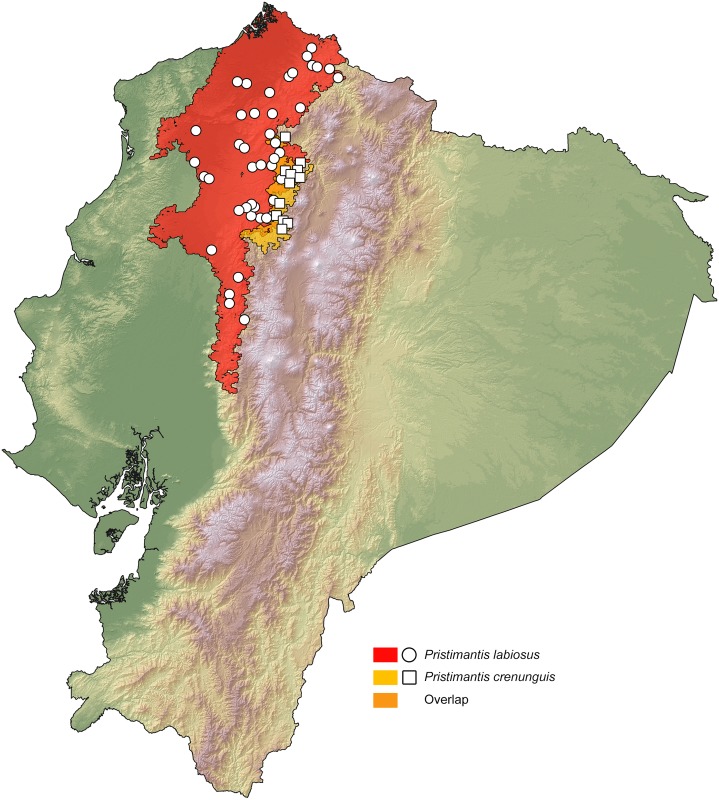
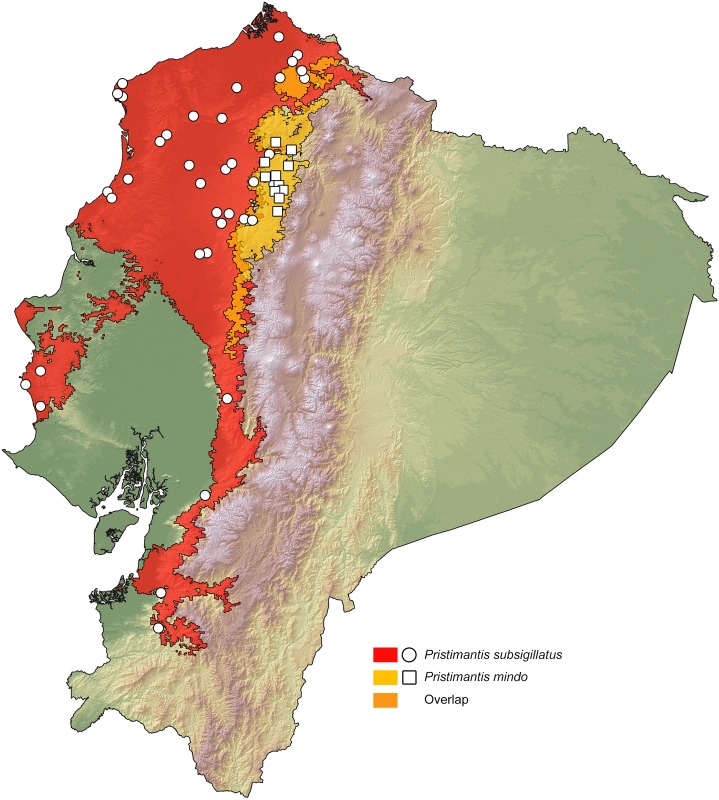
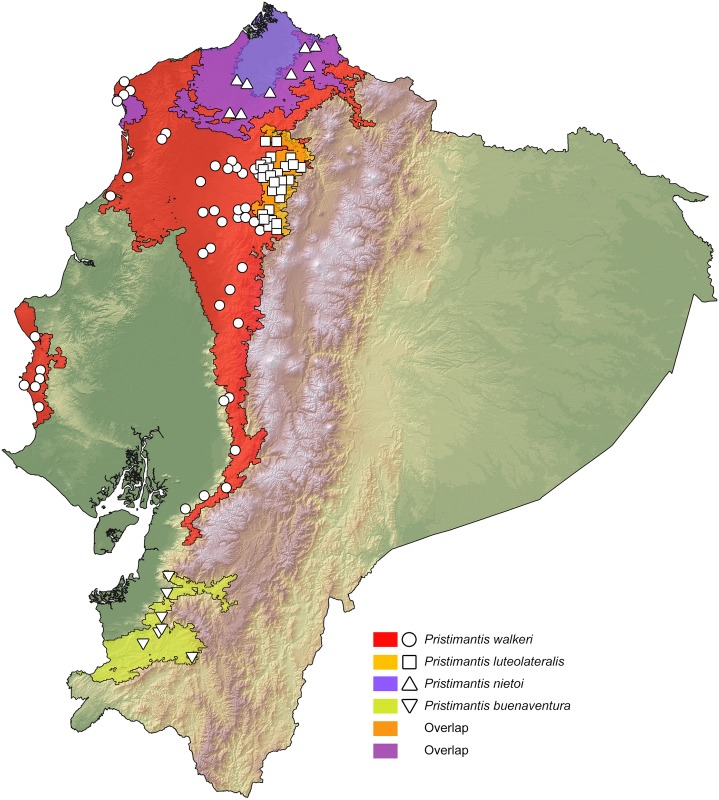
Comparative phylogeography allow us to understand how shared historical circumstances have shaped the formation of lineages, by examining a broad spectrum of co-distributed populations of different taxa. However, these types of studies are scarce in the Neotropics, a region that is characterized by high diversity, complex geology, and poorly understood biogeography. Here, we investigate the diversification patterns of five lineages of amphibians and reptiles, co-distributed across the Choco and Andes ecoregions in northwestern Ecuador. Mitochondrial DNA and occurrence records were used to determine the degree of geographic genetic divergence within species. Our results highlight congruent patterns of parapatric speciation and common geographical barriers for distantly related taxa. These comparisons indicate similar biological and demographic characteristics for the included clades, and reveal the existence of two new species of Pristimantis previously subsumed under P. walkeri, which we describe herein. Our data supports the hypothesis that widely distributed Chocoan taxa may generally experience their greatest opportunities for isolation and parapatric speciation across thermal elevational gradients. Finally, our study provides critical information to predict which unstudied lineages may harbor cryptic diversity, and how geology and climate are likely to have shaped their evolutionary history.
Conflict of interest statement
Figures










References
-
- Kessler M, Kluge J. Diversity and endemism in tropical montane forests—from patterns to processes In Gradstein SR, Homeier J, Gansert D, editors. The Tropical mountain forest: patterns and processes in a biodiversity hotspot. Göttingen: Göttingen Centre for Biodiversity and Ecology; 2008. pp. 35–50.
-
- Mittermeier RA, Turner WR, Larsen FW, Brooks TM, Gascon C. Global biodiversity conservation: the critical role of hotspots In Zachos FE, Hable JC, editors. Biodiversity hotspots. Heidelberg: Springer; 2011. pp. 3–22.
-
- Arteaga A, Bustamante L, Guayasamin JM. The amphibians and reptiles of Mindo. Quito: Universidad Tecnológica Indoamérica; 2013.
-
- Ortega-Andrade HM, Bermingham J, Aulestia C, Paucar C. Herpetofauna of the Bilsa Biological Station, province of Esmeraldas, Ecuador. Check List. 2010;6: 119–154.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
