

Topology of ON and OFF inputs in visual cortex enables an invariant columnar architecture
- PMID: 27120162
- PMCID: PMC5350615
- DOI: 10.1038/nature17941
Topology of ON and OFF inputs in visual cortex enables an invariant columnar architecture
Abstract
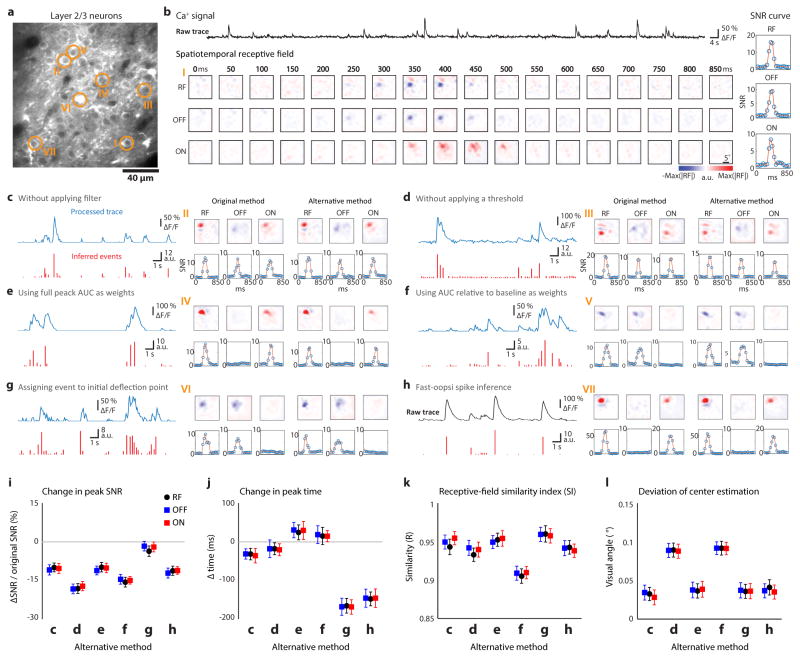
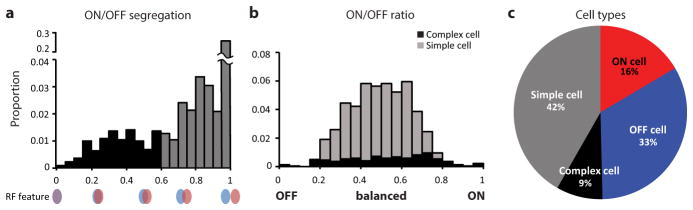
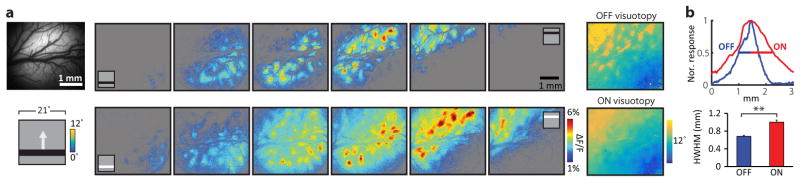
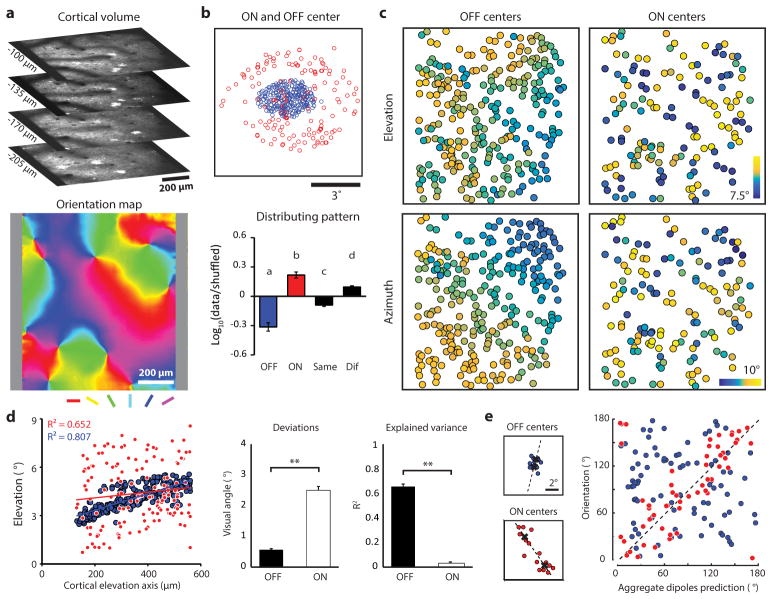
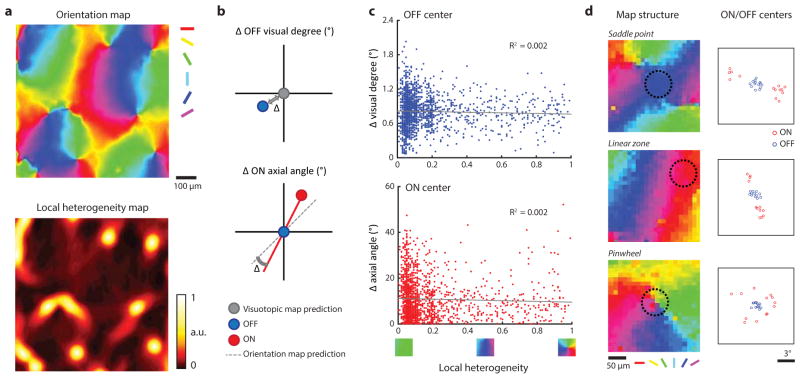
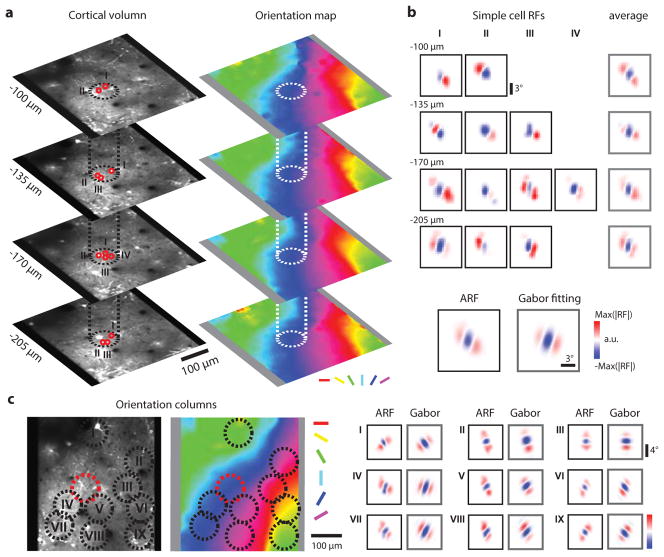
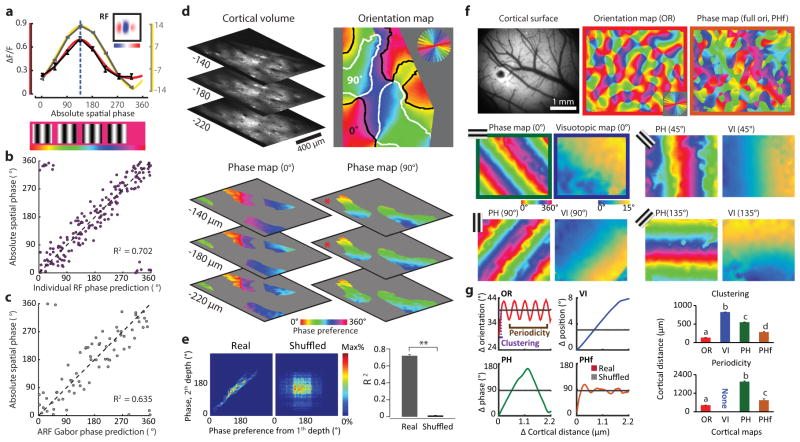
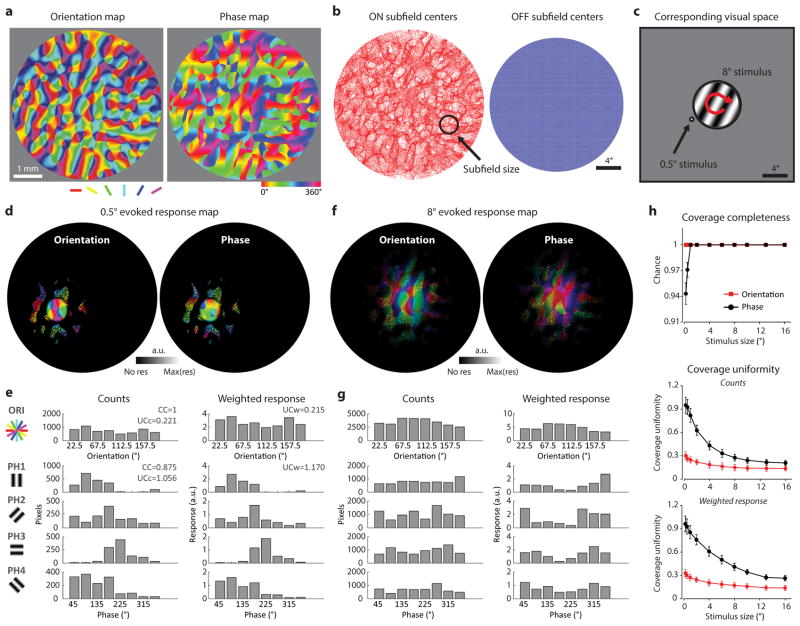
Circuits in the visual cortex integrate the information derived from separate ON (light-responsive) and OFF (dark-responsive) pathways to construct orderly columnar representations of stimulus orientation and visual space. How this transformation is achieved to meet the specific topographic constraints of each representation remains unclear. Here we report several novel features of ON-OFF convergence visualized by mapping the receptive fields of layer 2/3 neurons in the tree shrew (Tupaia belangeri) visual cortex using two-photon imaging of GCaMP6 calcium signals. We show that the spatially separate ON and OFF subfields of simple cells in layer 2/3 exhibit topologically distinct relationships with the maps of visual space and orientation preference. The centres of OFF subfields for neurons in a given region of cortex are confined to a compact region of visual space and display a smooth visuotopic progression. By contrast, the centres of the ON subfields are distributed over a wider region of visual space, display substantial visuotopic scatter, and have an orientation-specific displacement consistent with orientation preference map structure. As a result, cortical columns exhibit an invariant aggregate receptive field structure: an OFF-dominated central region flanked by ON-dominated subfields. This distinct arrangement of ON and OFF inputs enables continuity in the mapping of both orientation and visual space and the generation of a columnar map of absolute spatial phase.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Ferster D, Chung S, Wheat H. Orientation selectivity of thalamic input to simple cells of cat visual cortex. Nature. 1996;380:249–252. - PubMed
-
- Jin J, Wang Y, Swadlow HA, Alonso JM. Population receptive fields of ON and OFF thalamic inputs to an orientation column in visual cortex. Nat Neurosci. 2011;14:232–238. - PubMed
-
- Bosking WH, Crowley JC, Fitzpatrick D. Spatial coding of position and orientation in primary visual cortex. Nat Neurosci. 2002;5:874–882. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
