

Improved analysis of C4 and C3 photosynthesis via refined in vitro assays of their carbon fixation biochemistry
- PMID: 27122573
- PMCID: PMC4867899
- DOI: 10.1093/jxb/erw154
Improved analysis of C4 and C3 photosynthesis via refined in vitro assays of their carbon fixation biochemistry
Abstract
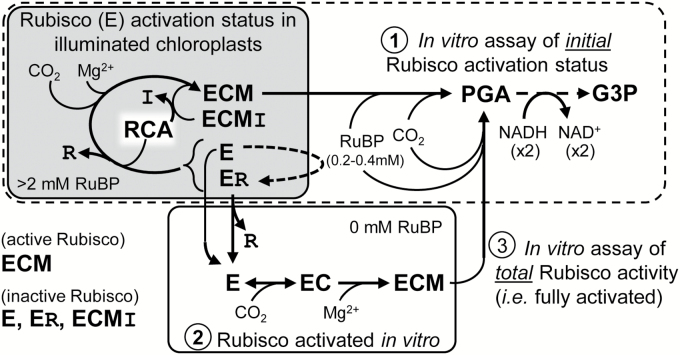
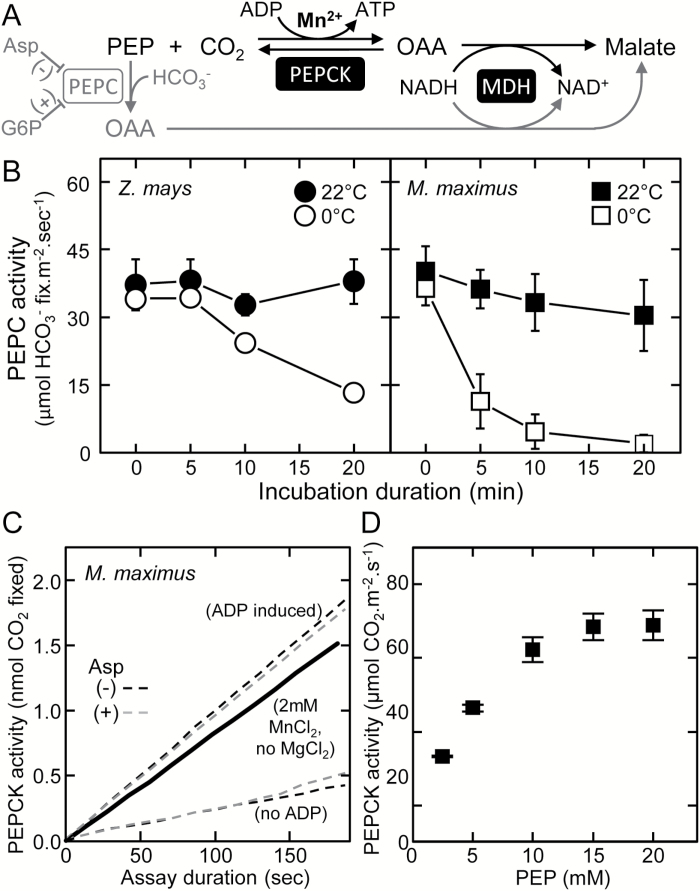
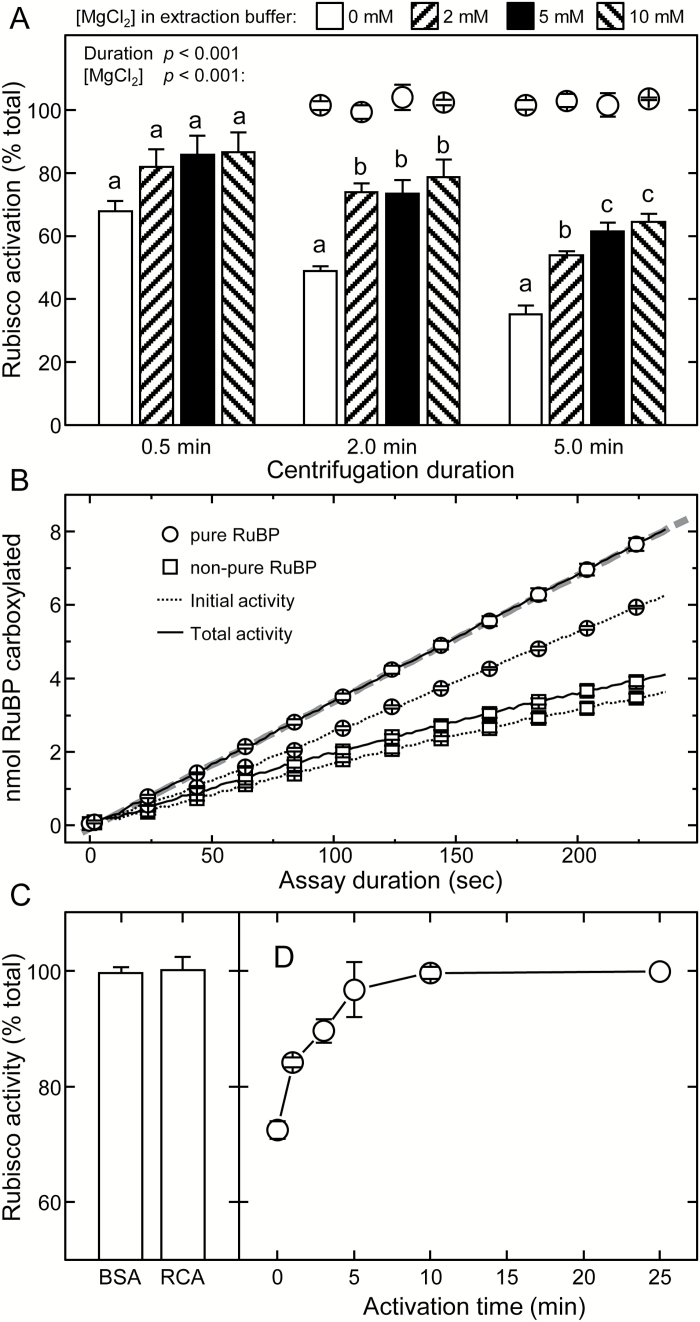
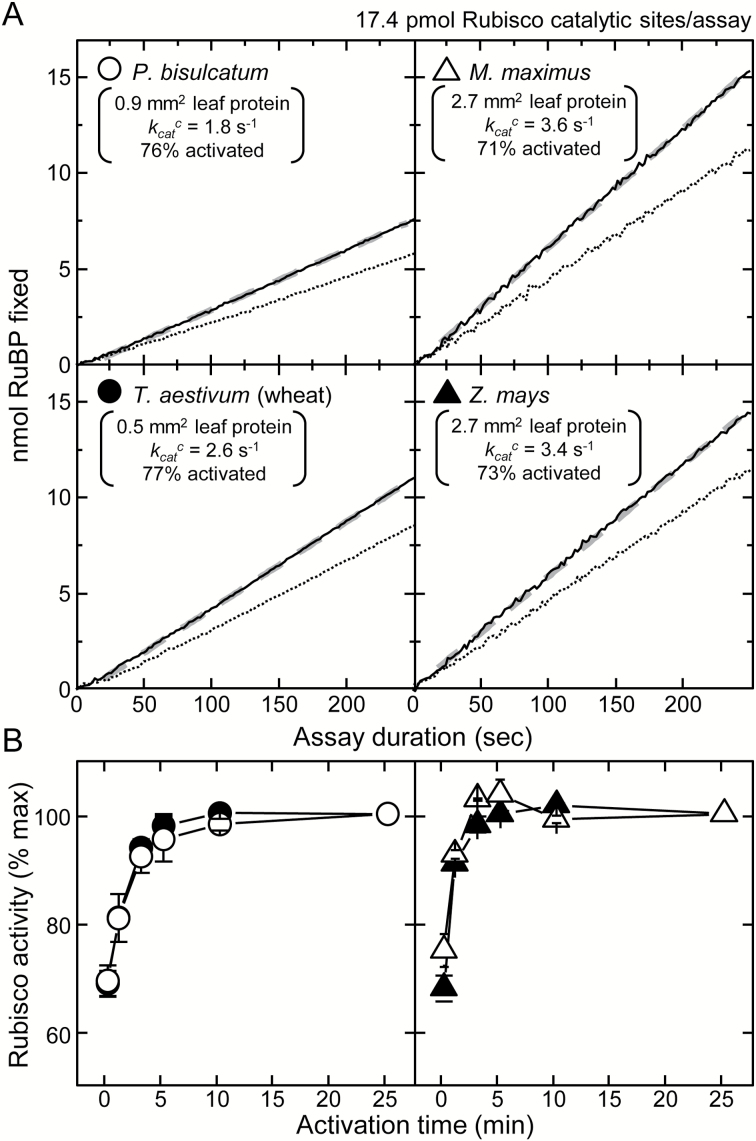
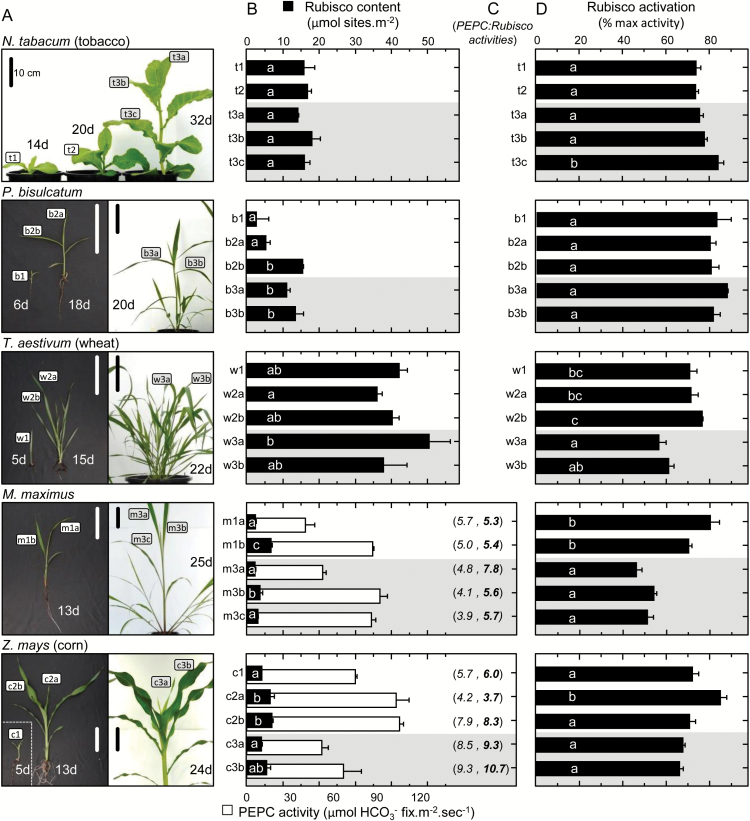
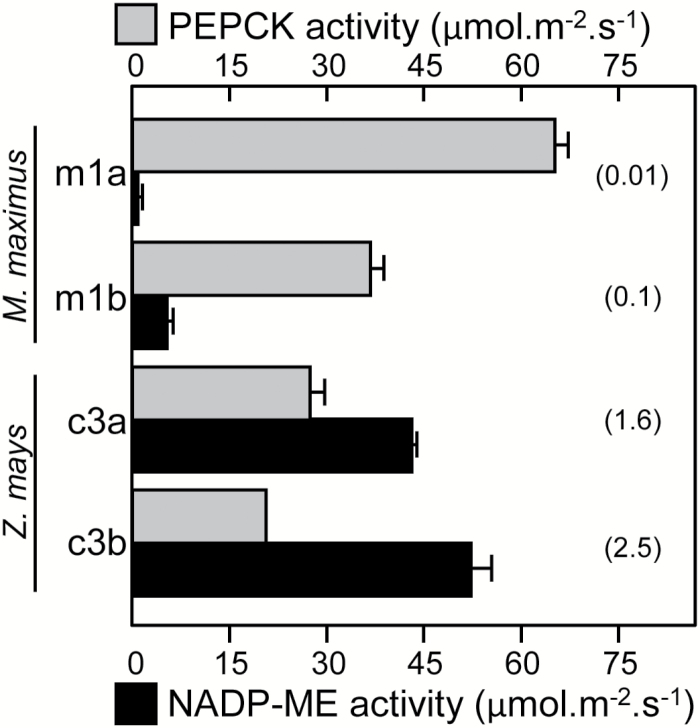
Plants operating C3 and C4 photosynthetic pathways exhibit differences in leaf anatomy and photosynthetic carbon fixation biochemistry. Fully understanding this underpinning biochemical variation is requisite to identifying solutions for improving photosynthetic efficiency and growth. Here we refine assay methods for accurately measuring the carboxylase and decarboxylase activities in C3 and C4 plant soluble protein. We show that differences in plant extract preparation and assay conditions are required to measure NADP-malic enzyme and phosphoenolpyruvate carboxylase (pH 8, Mg(2+), 22 °C) and phosphoenolpyruvate carboxykinase (pH 7, >2mM Mn(2+), no Mg(2+)) maximal activities accurately. We validate how the omission of MgCl2 during leaf protein extraction, lengthy (>1min) centrifugation times, and the use of non-pure ribulose-1,5-bisphosphate (RuBP) significantly underestimate Rubisco activation status. We show how Rubisco activation status varies with leaf ontogeny and is generally lower in mature C4 monocot leaves (45-60% activation) relative to C3 monocots (55-90% activation). Consistent with their >3-fold lower Rubisco contents, full Rubisco activation in soluble protein from C4 leaves (<5min) was faster than in C3 plant samples (<10min), with addition of Rubisco activase not required for full activation. We conclude that Rubisco inactivation in illuminated leaves primarily stems from RuBP binding to non-carbamylated enzyme, a state readily reversible by dilution during cellular protein extraction.
Keywords: CO2-concentrating mechanism; Carbamylation; Rubisco; Rubisco activase.; carbon fixation; photosynthesis.
© The Author 2016. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Andersson I. 2008. Catalysis and regulation in Rubisco. Journal of Experimental Botany 59, 1555–1568. - PubMed
-
- Andersson I, Backlund A. 2008. Structure and function of Rubisco. Plant Physiology and Biochemistry 46, 275–291. - PubMed
-
- Andralojc PJ, Madgwick PJ, Tao Y, et al. 2012. 2-Carboxy-d-arabinitol 1-phosphate (CA1P) phosphatase: evidence for a wider role in plant Rubisco regulation. Biochemical Journal 442, 733–742. - PubMed
-
- Andrews TJ, Whitney SM. 2003. Manipulating ribulose bisphosphate carboxylase/oxygenase in the chloroplasts of higher plants. Archives of Biochemistry and Biophysics 414, 159–169. - PubMed
-
- Anwaruzzaman, Nakano Y, Yokota A. 1996. Different location in dark-adapted leaves of Phaseolus vulgaris of ribulose-1,5-bisphosphate carboxylase/oxygenase and 2-carboxyarabinitol 1-phosphate. FEBS Letters 388, 223–227. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
