

CDKN2A-independent role of BMI1 in promoting growth and survival of Ph+ acute lymphoblastic leukemia
- PMID: 27125204
- PMCID: PMC4972639
- DOI: 10.1038/leu.2016.70
CDKN2A-independent role of BMI1 in promoting growth and survival of Ph+ acute lymphoblastic leukemia
Abstract
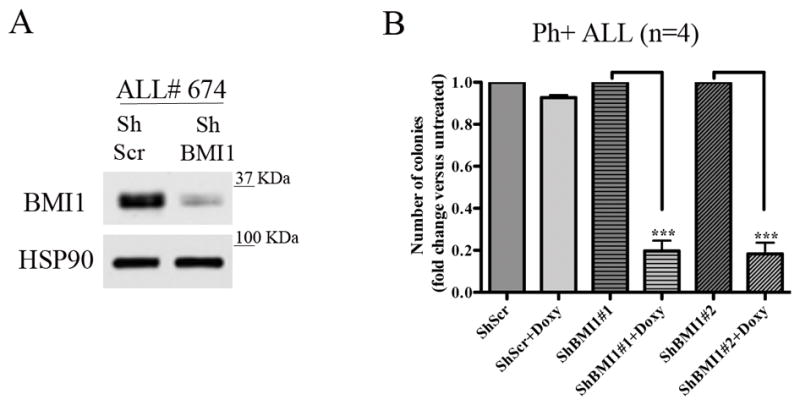
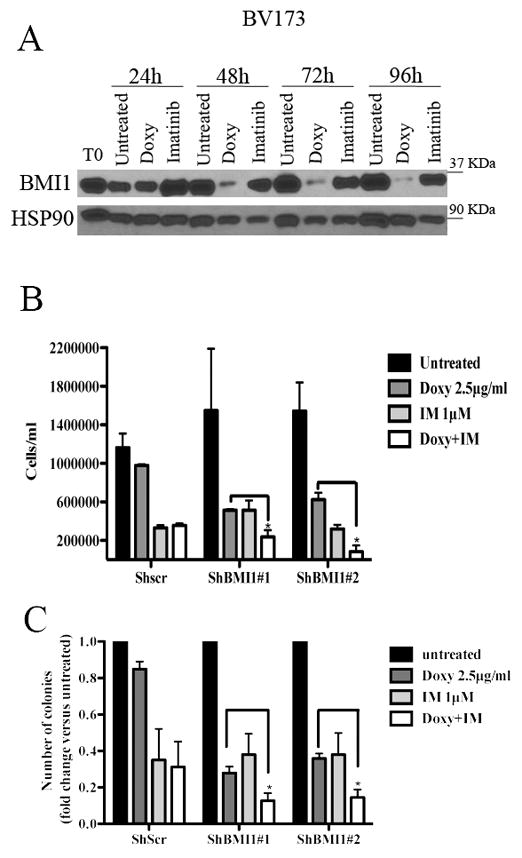
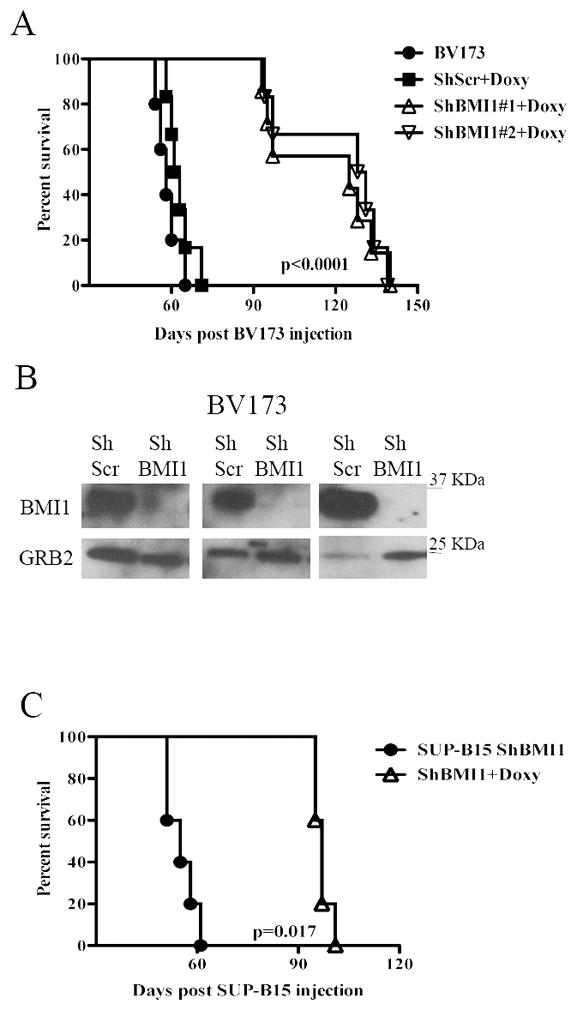
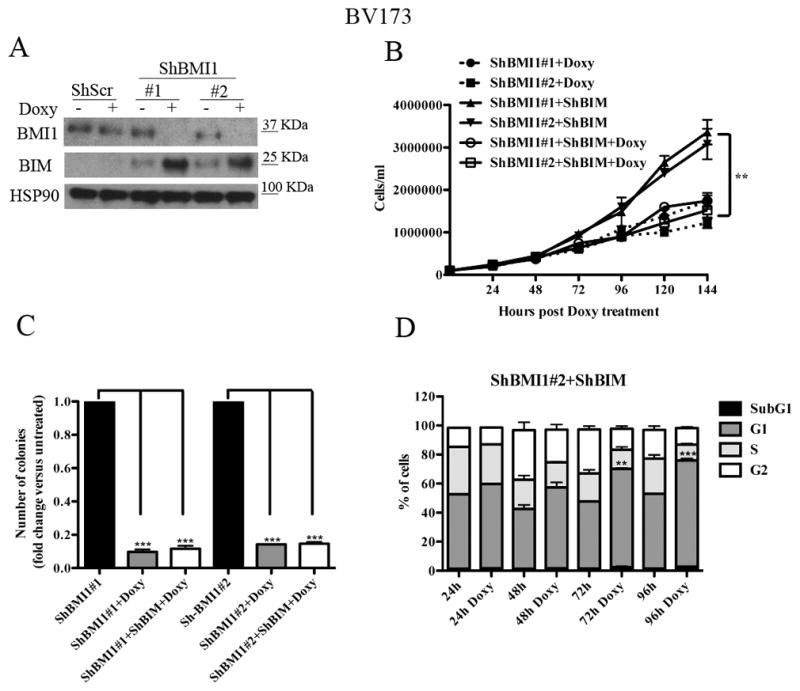
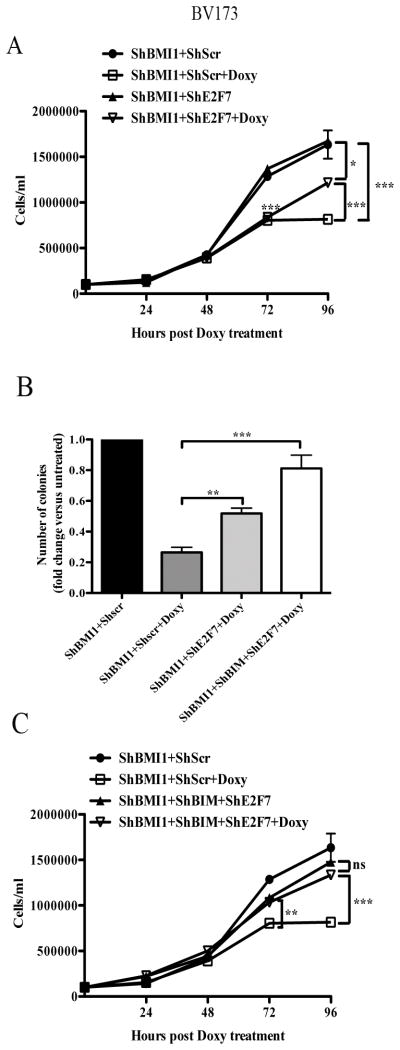
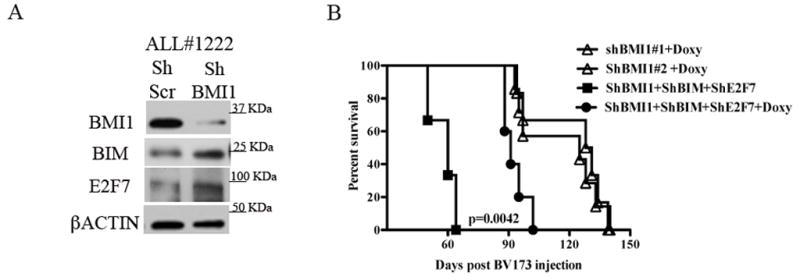
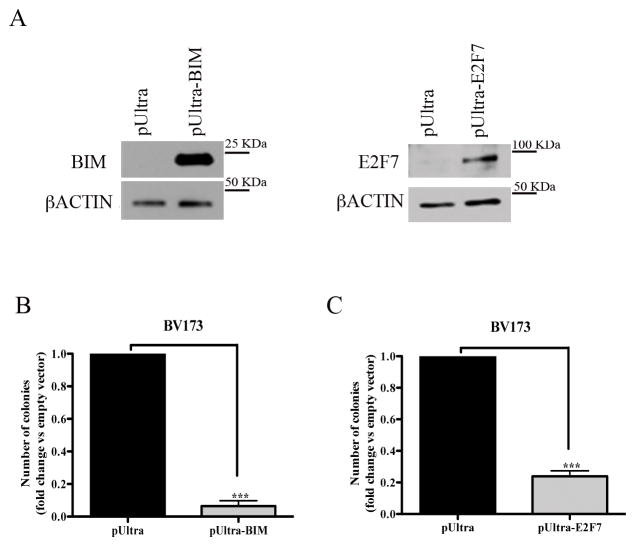
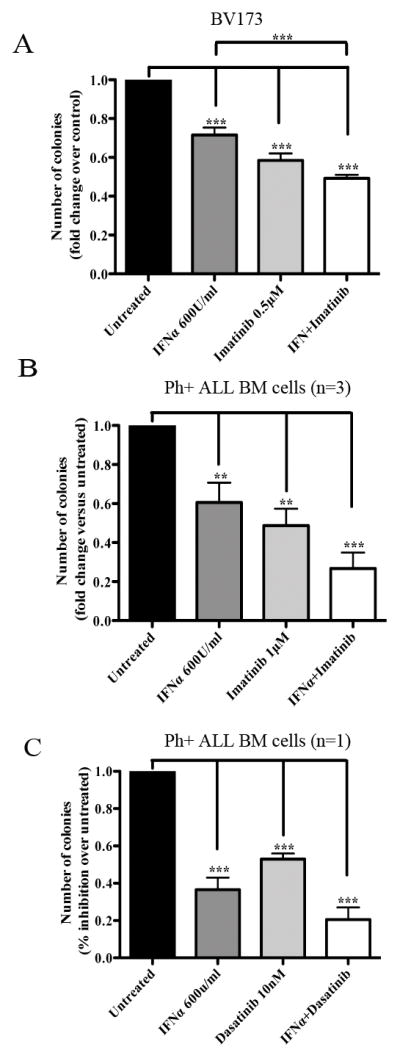
BMI1 is a key component of the PRC1 (polycomb repressive complex-1) complex required for maintenance of normal and cancer stem cells. Its aberrant expression is detected in chronic myeloid leukemia and Ph+ acute lymphoblastic leukemia (ALL), but no data exist on BMI1 requirement in ALL cells. We show here that BMI1 expression is important for proliferation and survival of Ph+ ALL cells and for leukemogenesis of Ph+ cells in vivo. Levels of BIM, interferon-α (IFNα)-regulated genes and E2F7 were upregulated in BMI1-silenced cells, suggesting that repressing their expression is important for BMI1 biological effects. Consistent with this hypothesis, we found that: (i) downregulation of BIM or E2F7 abrogated apoptosis or rescued, in part, the reduced proliferation and colony formation of BMI1 silenced BV173 cells; (ii) BIM/E2F7 double silencing further enhanced colony formation and in vivo leukemogenesis of BMI1-silenced cells; (iii) overexpression of BIM and E2F7 mimicked the effect of BMI1 silencing in BV173 and SUP-B15 cells; and (iv) treatment with IFNα suppressed proliferation and colony formation of Ph+ ALL cells. These studies indicate that the growth-promoting effects of BMI1 in Ph+ ALL cells depend on suppression of multiple pathways and support the use of IFNα in the therapy of Ph+ ALL.
Conflict of interest statement
The authors declare no conflict of interest
Figures
References
-
- Melo JV. The diversity of BCR-ABL fusion proteins and their relationShip to leukemia phenotype. Blood. 1996;88:2375–2384. - PubMed
-
- Goga A, McLaughlin J, Afar DE, Saffran DC, Witte ON. Alternative signals to RAS for hematopoietic transformation by the BCR-ABL oncogene. Cell. 1995;82:981–988. - PubMed
-
- Nieborowska-Skorska M, Wasik MA, Slupianek A, Salomoni P, Kitamura T, Calabretta B, et al. Signal transducer and activator of transcription (STAT)5 activation by BCR/ABLis dependent on intact Src homology (SH)3 and SH2 domains of BCR/ABL and is required for leukemogenesis. J Exp Med. 1999;189:1229–1242. - PMC - PubMed
-
- Perrotti D, Calabretta B. The biology of CML blast crisis. Blood. 2004;103:4010–4022. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
