

How Aromatic Compounds Block DNA Binding of HcaR Catabolite Regulator
- PMID: 27129205
- PMCID: PMC4933237
- DOI: 10.1074/jbc.M115.712067
How Aromatic Compounds Block DNA Binding of HcaR Catabolite Regulator
Abstract
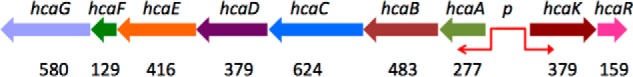
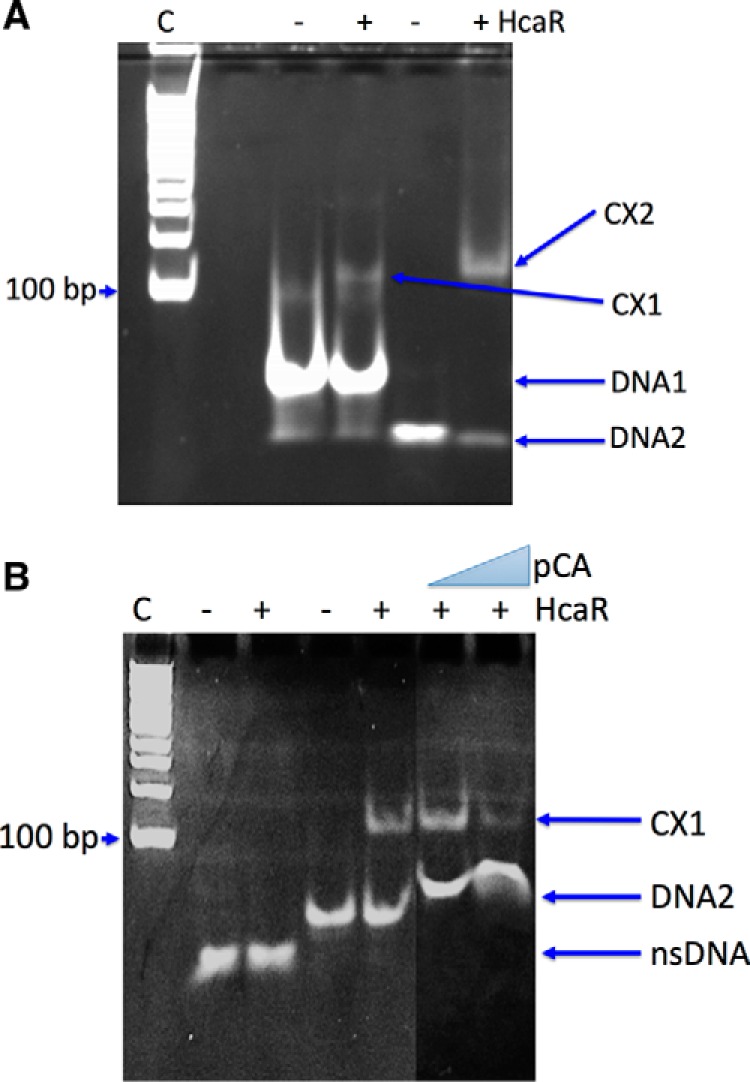
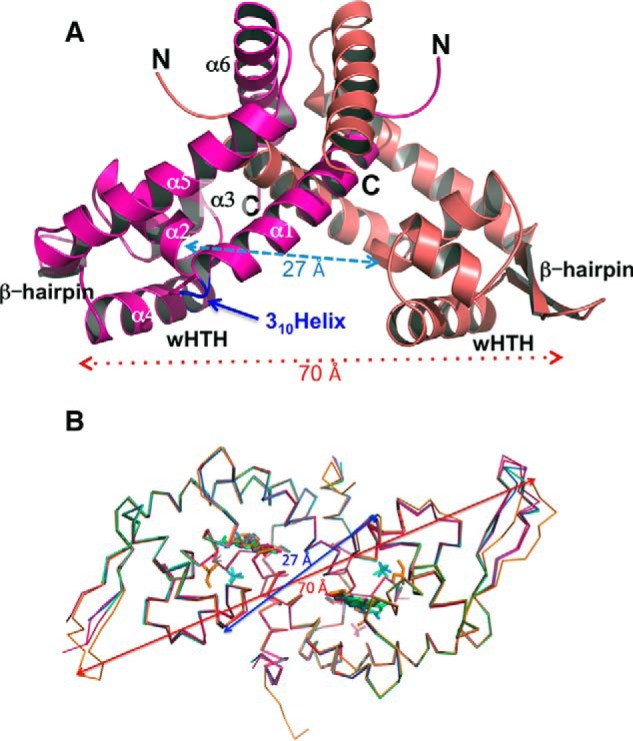
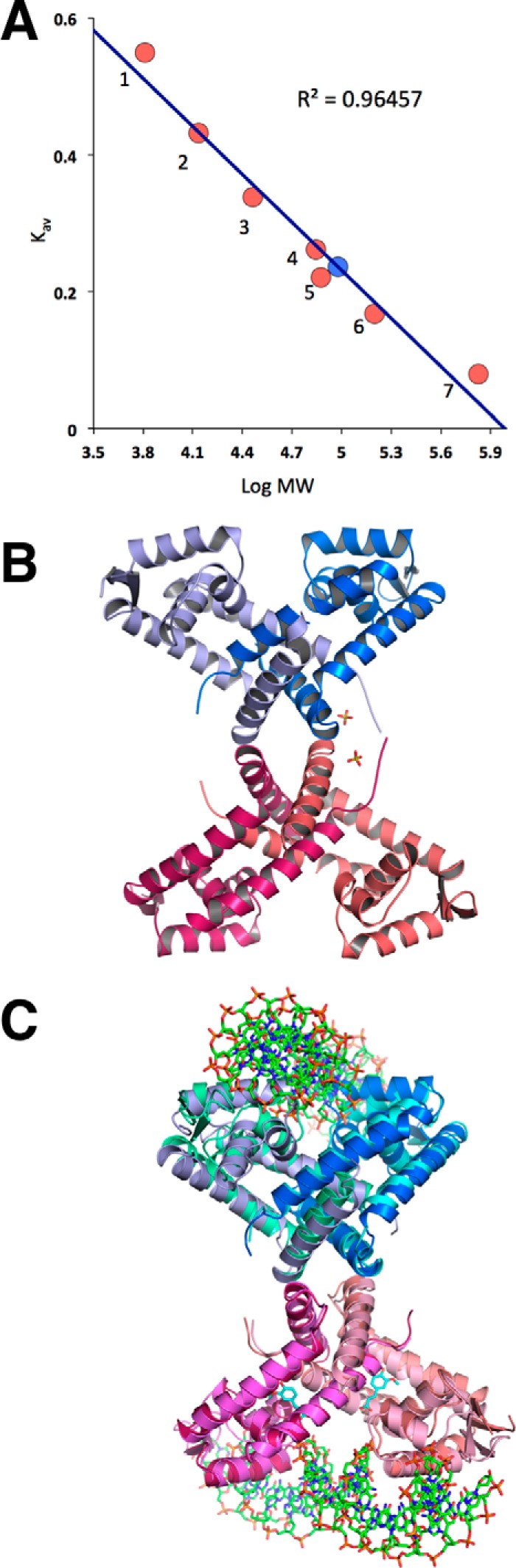
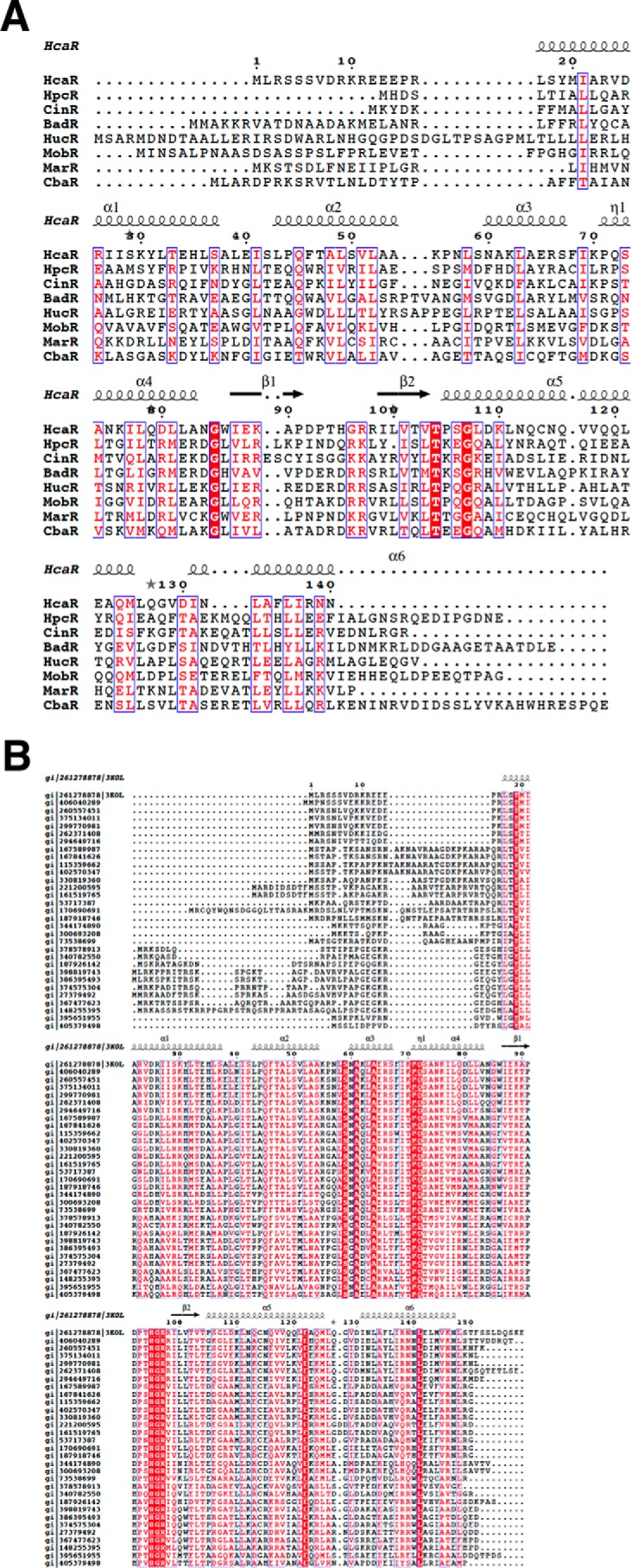
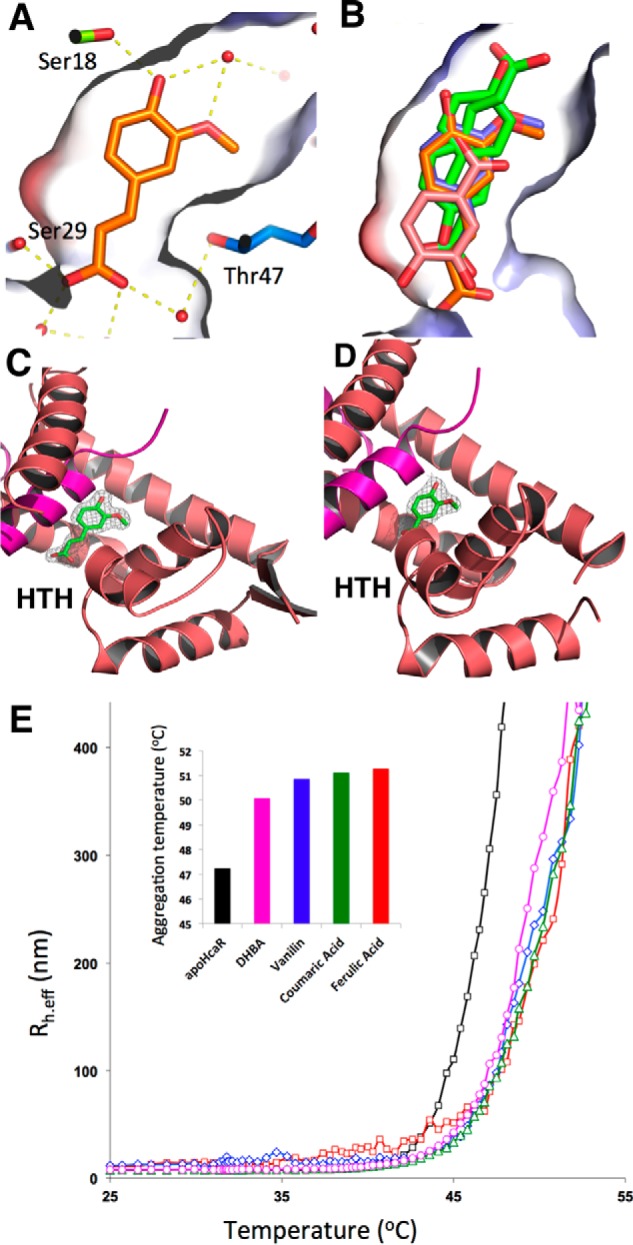
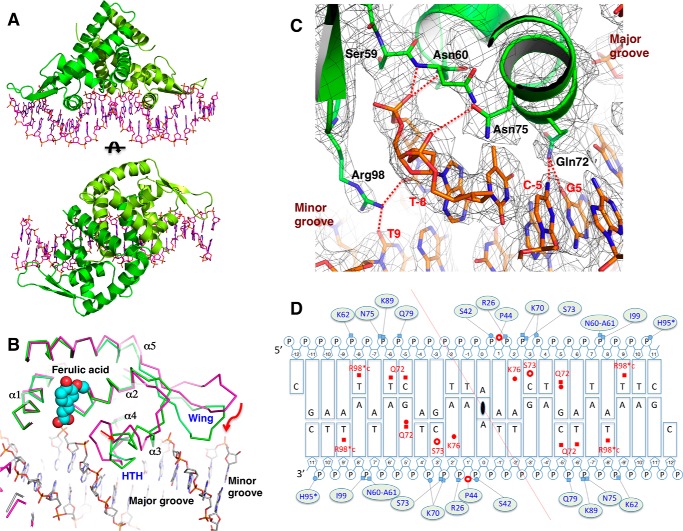
Bacterial catabolism of aromatic compounds from various sources including phenylpropanoids and flavonoids that are abundant in soil plays an important role in the recycling of carbon in the ecosystem. We have determined the crystal structures of apo-HcaR from Acinetobacter sp. ADP1, a MarR/SlyA transcription factor, in complexes with hydroxycinnamates and a specific DNA operator. The protein regulates the expression of the hca catabolic operon in Acinetobacter and related bacterial strains, allowing utilization of hydroxycinnamates as sole sources of carbon. HcaR binds multiple ligands, and as a result the transcription of genes encoding several catabolic enzymes is increased. The 1.9-2.4 Å resolution structures presented here explain how HcaR recognizes four ligands (ferulate, 3,4-dihydroxybenzoate, p-coumarate, and vanillin) using the same binding site. The ligand promiscuity appears to be an adaptation to match a broad specificity of hydroxycinnamate catabolic enzymes while responding to toxic thioester intermediates. Structures of apo-HcaR and in complex with a specific DNA hca operator when combined with binding studies of hydroxycinnamates show how aromatic ligands render HcaR unproductive in recognizing a specific DNA target. The current study contributes to a better understanding of the hca catabolic operon regulation mechanism by the transcription factor HcaR.
Keywords: DNA binding; DNA-binding protein; NUDIX family; arabinose utilization; bacterial transcription; human gut bacteria; ligand-binding protein; ligand-induced conformational change; metabolic regulation; protein conformation.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Schumacher M. A., and Brennan R. G. (2002) Structural mechanisms of multidrug recognition and regulation by bacterial multidrug transcription factors. Mol. Microbiol. 45, 885–893 - PubMed
-
- Alekshun M. N., and Levy S. B. (1999) The mar regulon: multiple resistance to antibiotics and other toxic chemicals. Trends Microbiol. 7, 410–413 - PubMed
-
- Davis J. R., and Sello J. K. (2010) Regulation of genes in Streptomyces bacteria required for catabolism of lignin-derived aromatic compounds. Appl. Microbiol. Biotechnol. 86, 921–929 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
