

Zinc Finger Homeodomain Factor Zfhx3 Is Essential for Mammary Lactogenic Differentiation by Maintaining Prolactin Signaling Activity
- PMID: 27129249
- PMCID: PMC4933452
- DOI: 10.1074/jbc.M116.719377
Zinc Finger Homeodomain Factor Zfhx3 Is Essential for Mammary Lactogenic Differentiation by Maintaining Prolactin Signaling Activity
Abstract
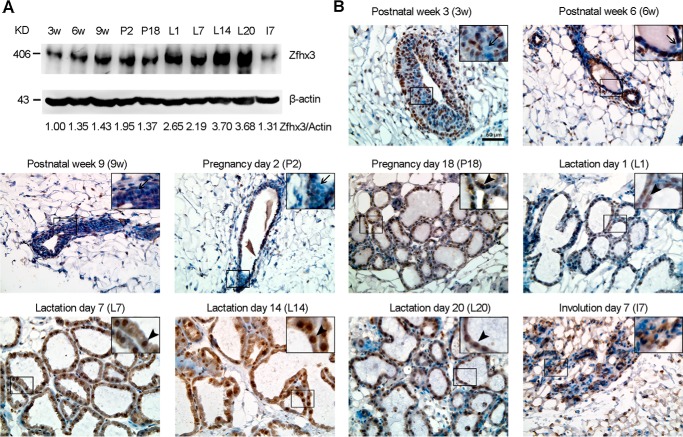
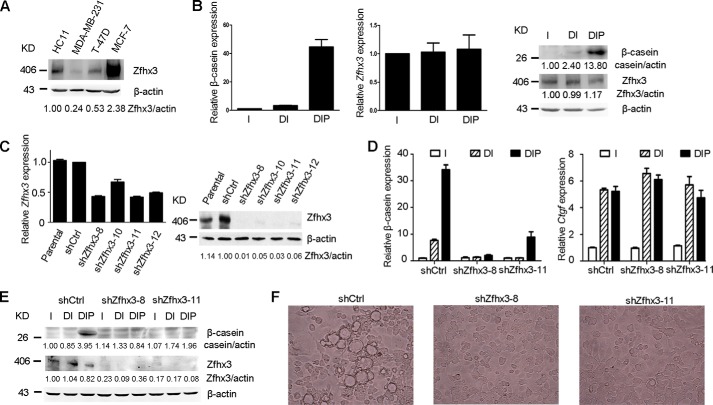
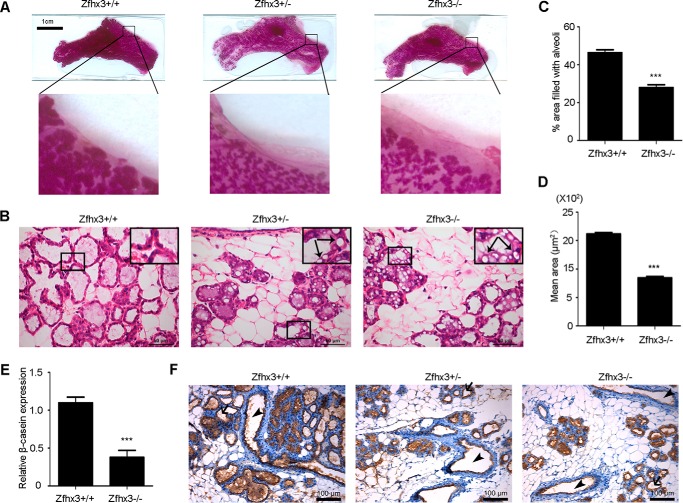
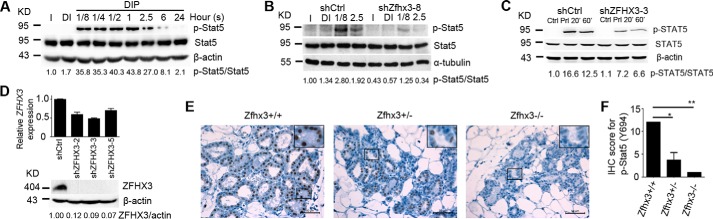
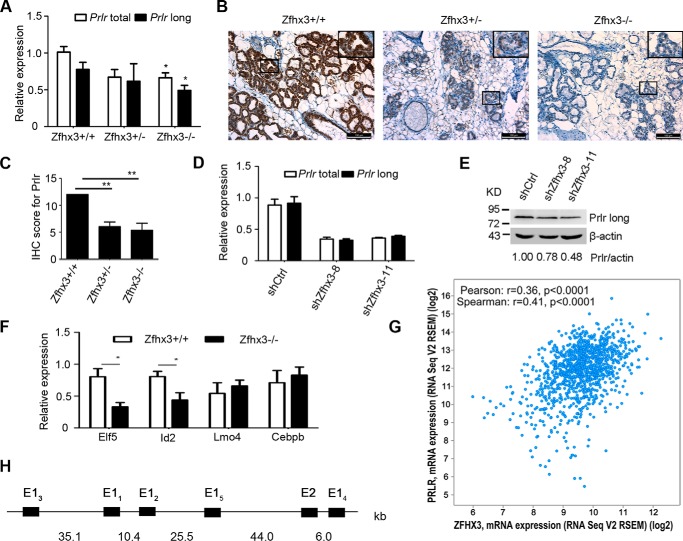
The zinc finger homeobox 3 (ZFHX3, also named ATBF1 for AT motif binding factor 1) is a transcription factor that suppresses prostatic carcinogenesis and induces neuronal differentiation. It also interacts with estrogen receptor α to inhibit cell proliferation and regulate pubertal mammary gland development in mice. In the present study, we examined whether and how Zfhx3 regulates lactogenic differentiation in mouse mammary glands. At different stages of mammary gland development, Zfhx3 protein was expressed at varying levels, with the highest level at lactation. In the HC11 mouse mammary epithelial cell line, an in vitro model of lactogenesis, knockdown of Zfhx3 attenuated prolactin-induced β-casein expression and morphological changes, indicators of lactogenic differentiation. In mouse mammary tissue, knock-out of Zfhx3 interrupted lactogenesis, resulting in underdeveloped glands with much smaller and fewer alveoli, reduced β-casein expression, accumulation of large cytoplasmic lipid droplets in luminal cells after parturition, and failure in lactation. Mechanistically, Zfhx3 maintained the expression of Prlr (prolactin receptor) and Prlr-Jak2-Stat5 signaling activity, whereas knockdown and knock-out of Zfhx3 in HC11 cells and mammary tissues, respectively, decreased Prlr expression, Stat5 phosphorylation, and the expression of Prlr-Jak2-Stat5 target genes. These findings indicate that Zfhx3 plays an essential role in proper lactogenic development in mammary glands, at least in part by maintaining Prlr expression and Prlr-Jak2-Stat5 signaling activity.
Keywords: STAT5 phosphorylation; ZFHX3/ATBF1; development; homeobox; lactogenic differentiation; mammary gland; peptide hormone; prolactin; prolactin receptor (PRLR); transcription factor.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Hennighausen L., and Robinson G. W. (2001) Signaling pathways in mammary gland development. Dev. Cell 1, 467–475 - PubMed
-
- Hennighausen L., and Robinson G. W. (2005) Information networks in the mammary gland. Nat. Rev. Mol. Cell Biol. 6, 715–725 - PubMed
-
- Silva C. M., and Shupnik M. A. (2007) Integration of steroid and growth factor pathways in breast cancer: focus on signal transducers and activators of transcription and their potential role in resistance. Mol. Endocrinol. 21, 1499–1512 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
