

The Dynamics and Turnover of Tau Aggregates in Cultured Cells: INSIGHTS INTO THERAPIES FOR TAUOPATHIES
- PMID: 27129267
- PMCID: PMC4933232
- DOI: 10.1074/jbc.M115.712083
The Dynamics and Turnover of Tau Aggregates in Cultured Cells: INSIGHTS INTO THERAPIES FOR TAUOPATHIES
Abstract
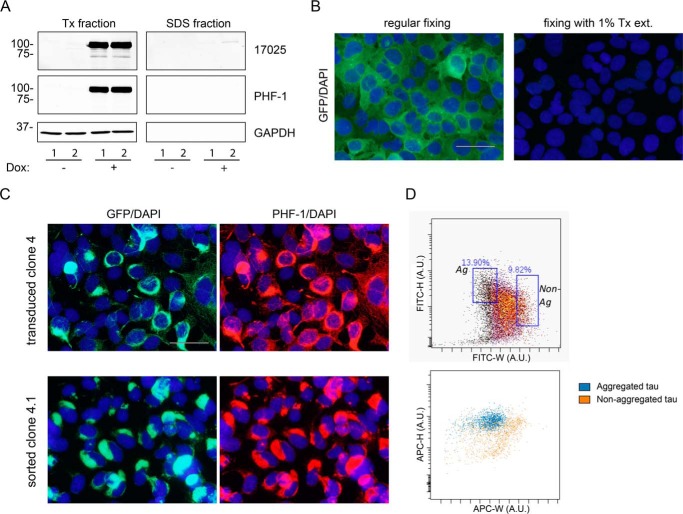
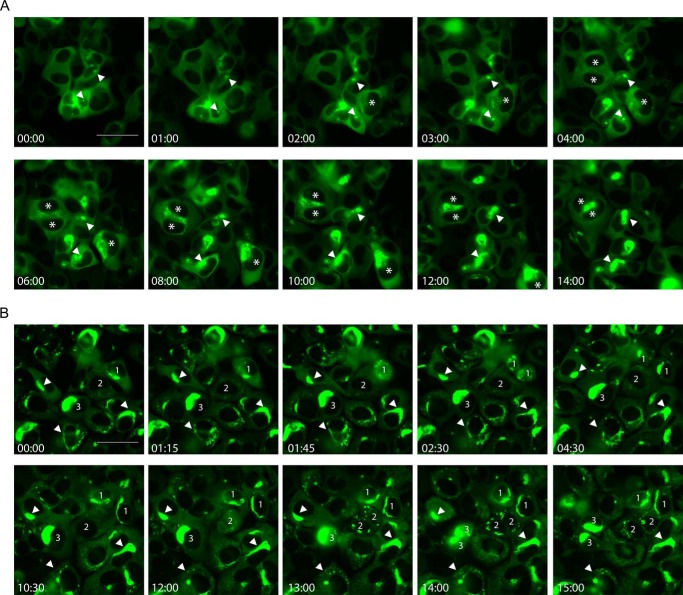
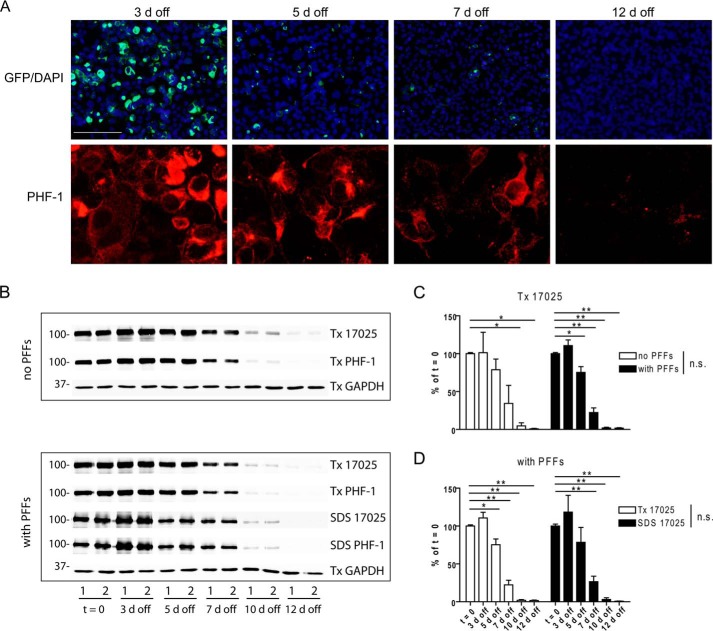
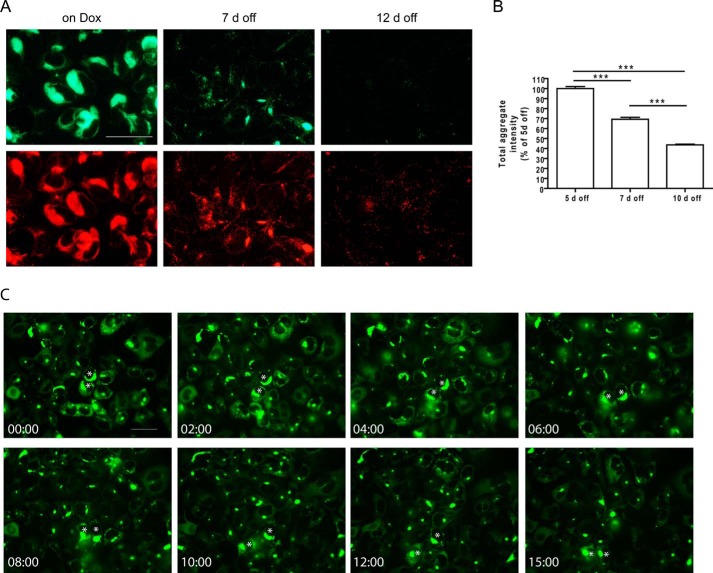
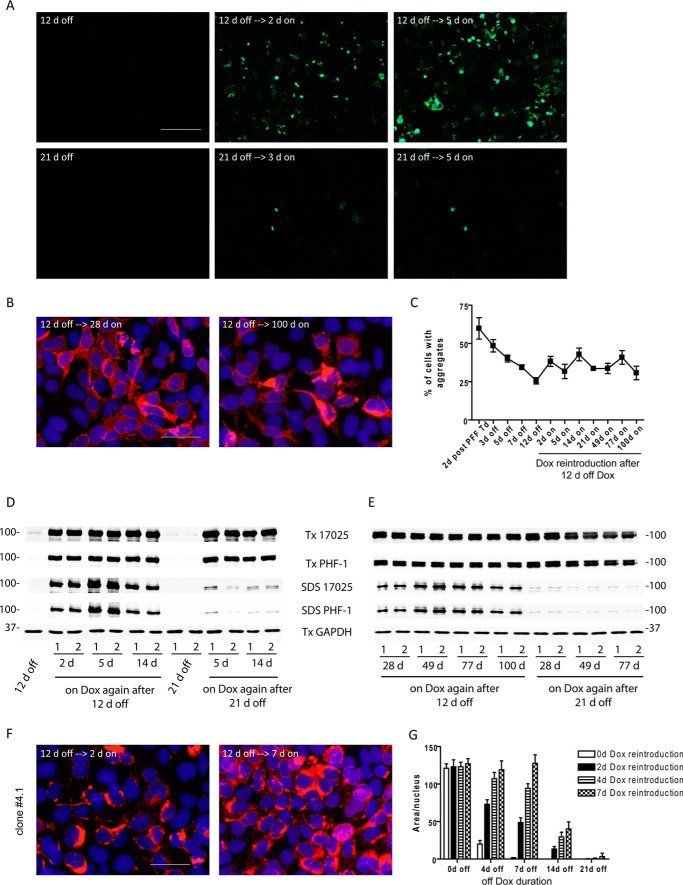
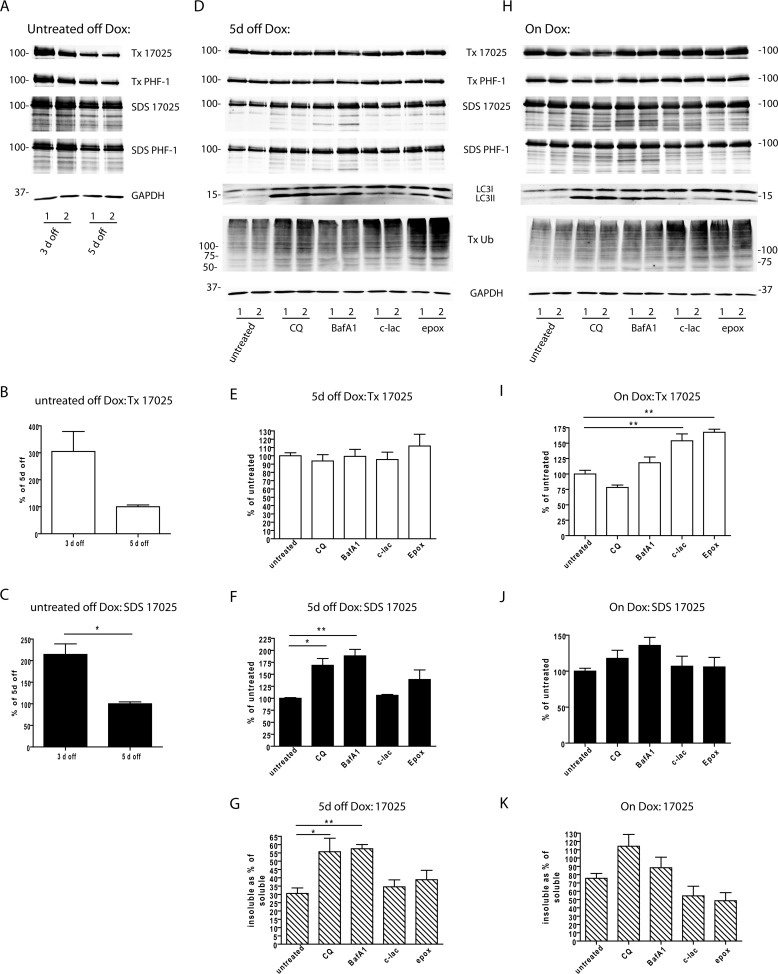
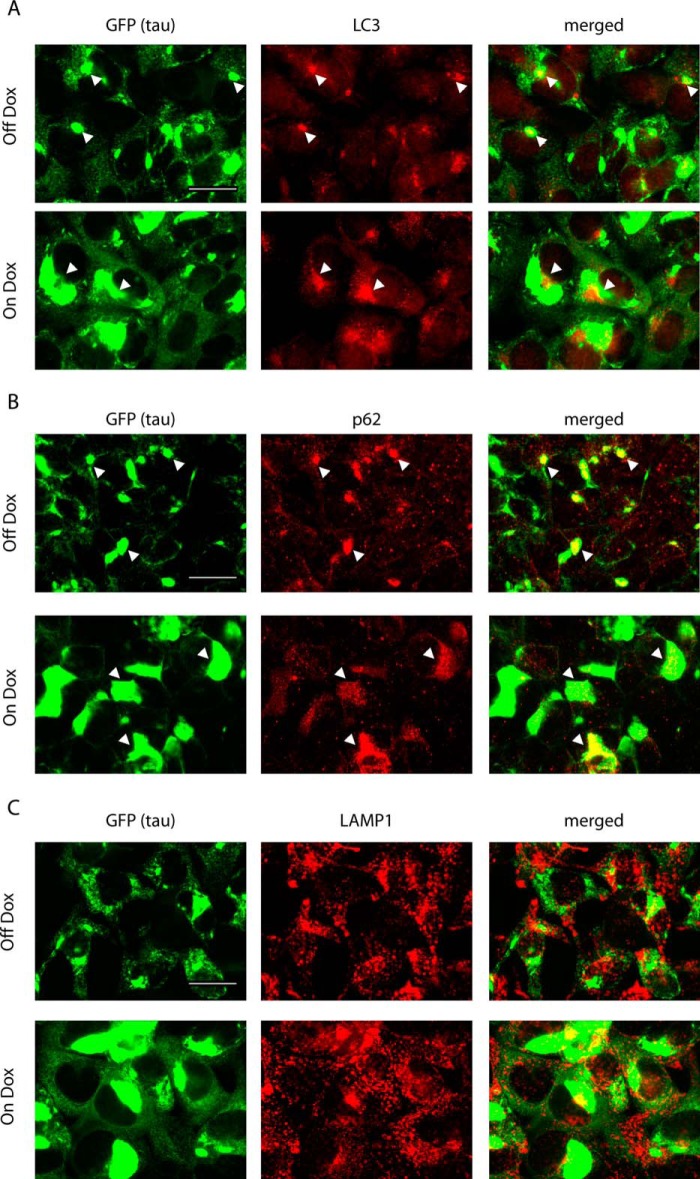
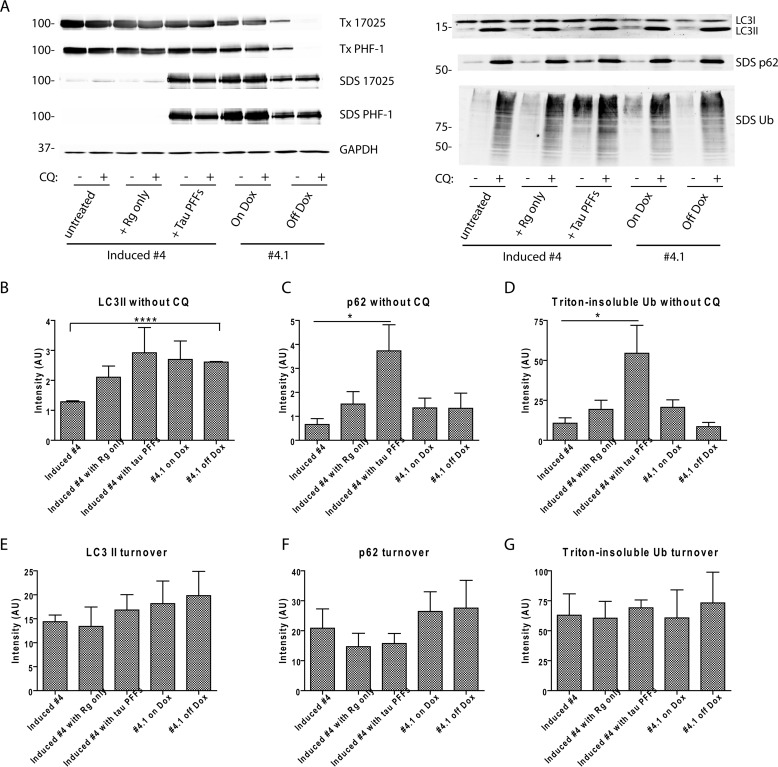
Filamentous tau aggregates, the hallmark lesions of Alzheimer disease (AD), play key roles in neurodegeneration. Activation of protein degradation systems has been proposed to be a potential strategy for removing pathological tau, but it remains unclear how effectively tau aggregates can be degraded by these systems. By applying our previously established cellular model system of AD-like tau aggregate induction using preformed tau fibrils, we demonstrate that tau aggregates induced in cells with regulated expression of full-length mutant tau can be gradually cleared when soluble tau expression is suppressed. This clearance is at least partially mediated by the autophagy-lysosome pathway, although both the ubiquitin-proteasome system and the autophagy-lysosome pathway are deficient in handling large tau aggregates. Importantly, residual tau aggregates left after the clearance phase leads to a rapid reinstatement of robust tau pathology once soluble tau expression is turned on again. Moreover, we succeeded in generating monoclonal cells persistently carrying tau aggregates without obvious cytotoxicity. Live imaging of GFP-tagged tau aggregates showed that tau inclusions are dynamic structures constantly undergoing "fission" and "fusion," which facilitate stable propagation of tau pathology in dividing cells. These findings provide a greater understanding of cell-to-cell transmission of tau aggregates in dividing cells and possibly neurons.
Keywords: Alzheimer disease; autophagy; protein aggregation; protein degradation; tau protein (tau).
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures


References
-
- Lee V. M., Goedert M., and Trojanowski J. Q. (2001) Neurodegenerative tauopathies. Annu. Rev. Neurosci. 24, 1121–1159 - PubMed
-
- Cleveland D. W., Hwo S. Y., and Kirschner M. W. (1977) Purification of tau, a microtubule-associated protein that induces assembly of microtubules from purified tubulin. J. Mol. Biol. 116, 207–225 - PubMed
-
- Ballatore C., Lee V. M., and Trojanowski J. Q. (2007) tau-mediated neurodegeneration in Alzheimer's disease and related disorders. Nat. Rev. Neurosci. 8, 663–672 - PubMed
-
- Arriagada P. V., Growdon J. H., Hedley-Whyte E. T., and Hyman B. T. (1992) Neurofibrillary tangles but not senile plaques parallel duration and severity of Alzheimer's disease. Neurology 42, 631–639 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
