

Chronic Activation of γ2 AMPK Induces Obesity and Reduces β Cell Function
- PMID: 27133129
- PMCID: PMC4873618
- DOI: 10.1016/j.cmet.2016.04.003
Chronic Activation of γ2 AMPK Induces Obesity and Reduces β Cell Function
Abstract
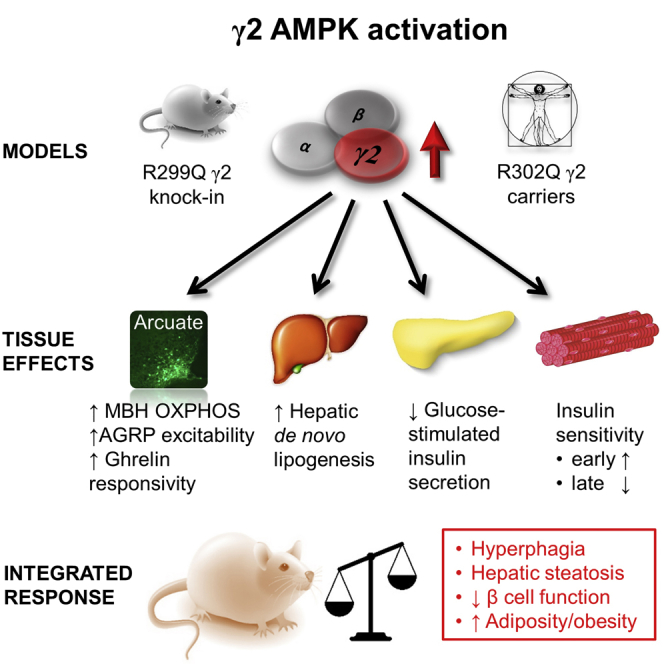
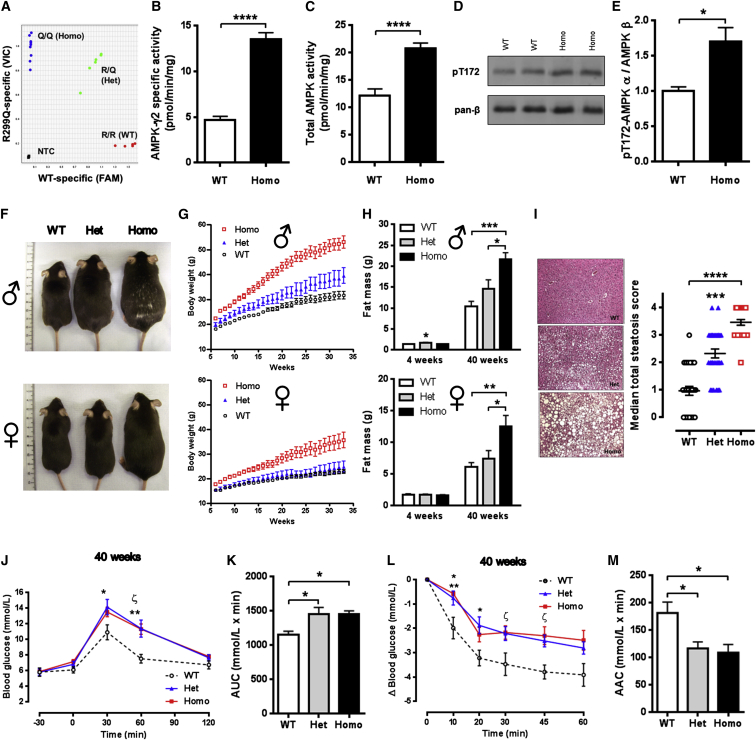
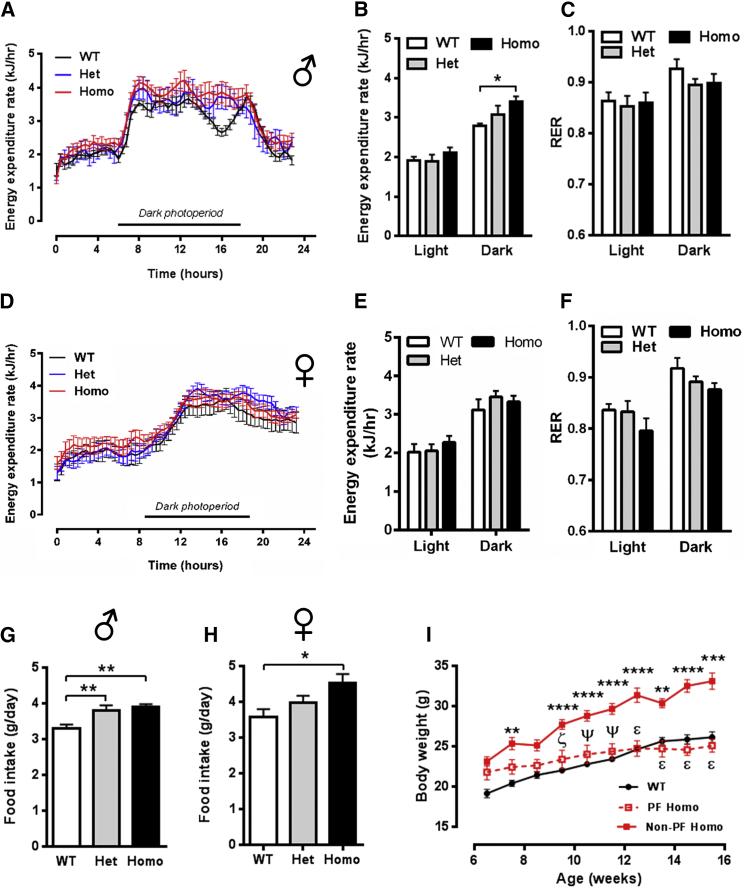
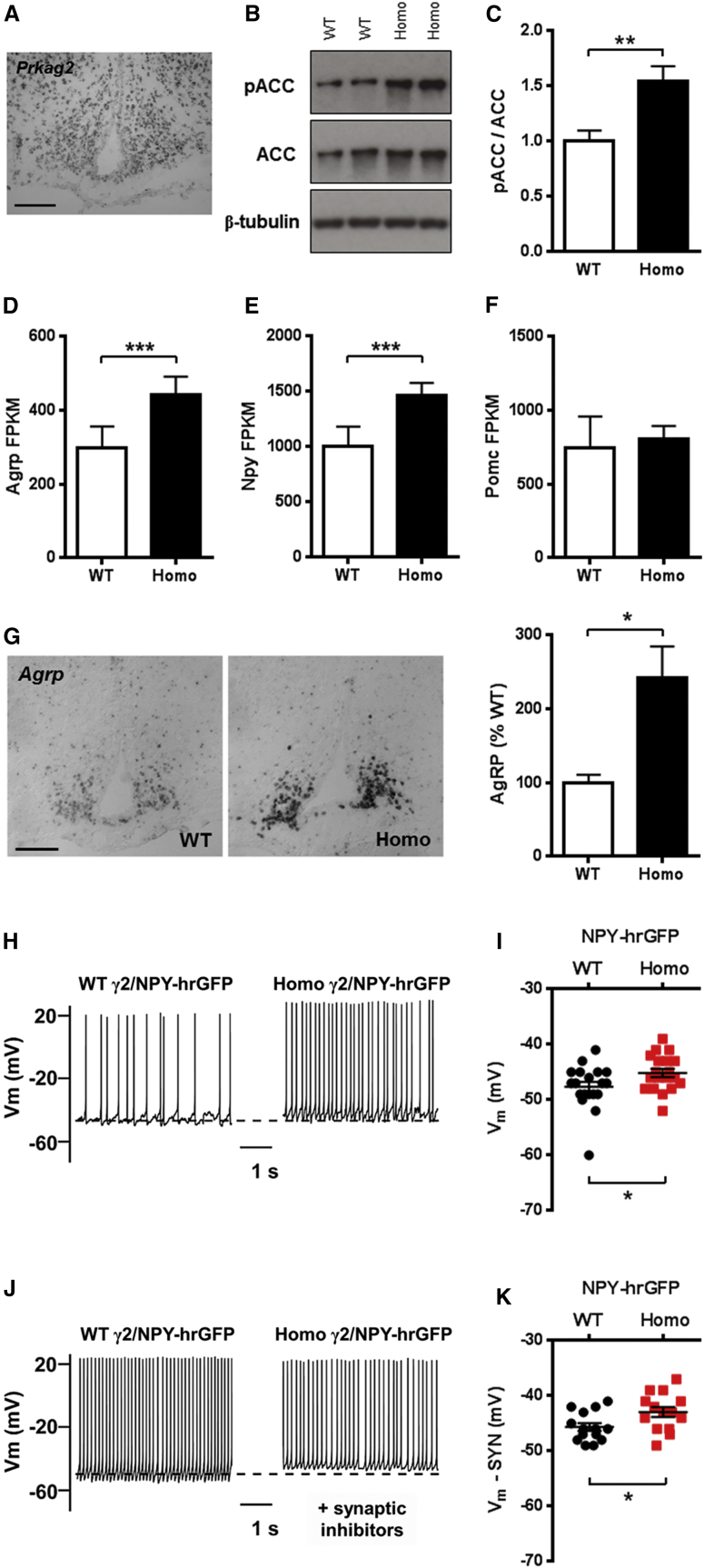
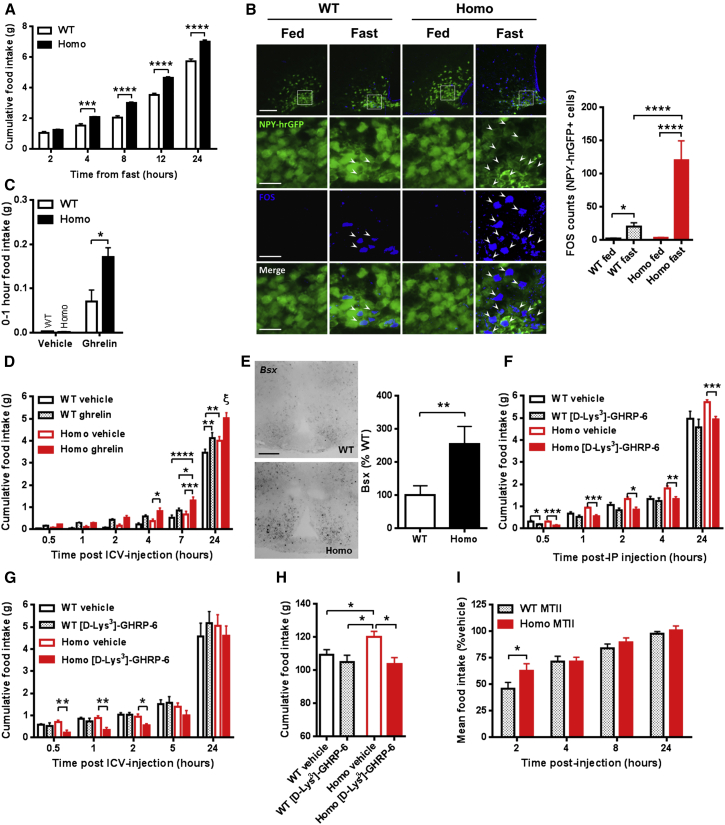
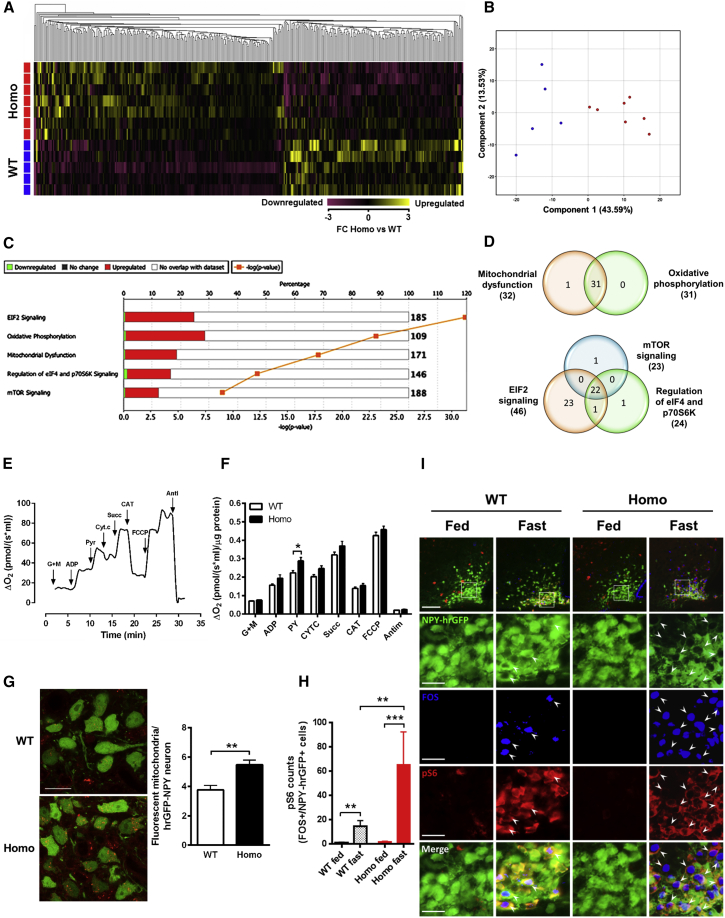
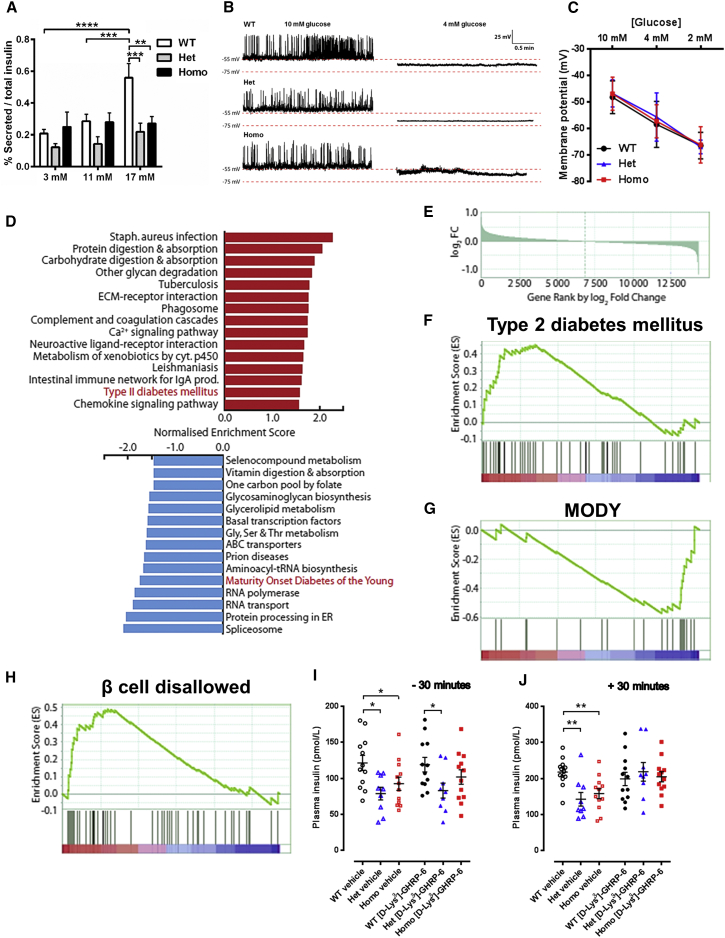
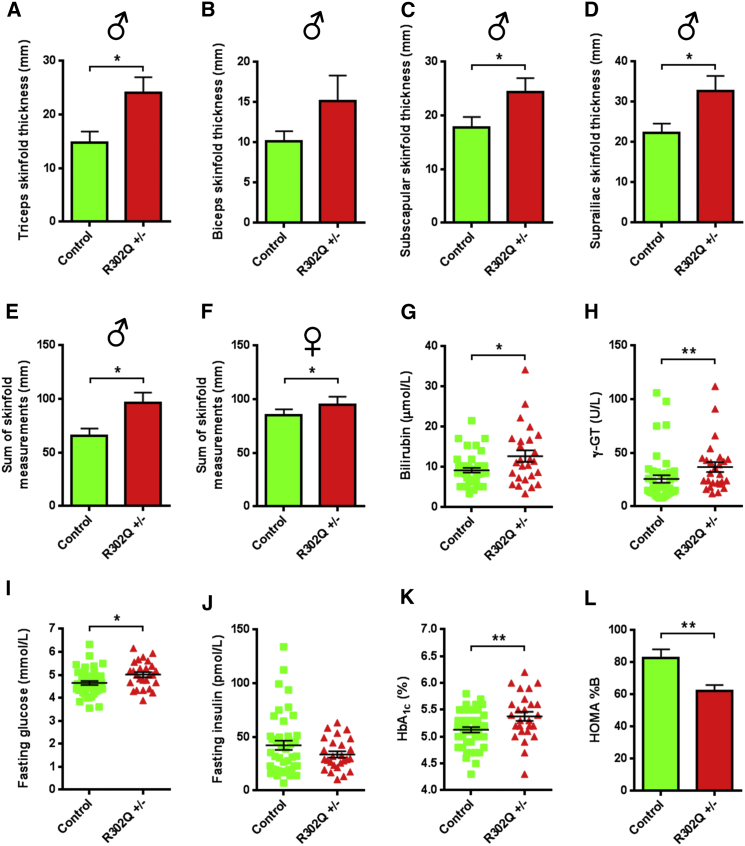
Despite significant advances in our understanding of the biology determining systemic energy homeostasis, the treatment of obesity remains a medical challenge. Activation of AMP-activated protein kinase (AMPK) has been proposed as an attractive strategy for the treatment of obesity and its complications. AMPK is a conserved, ubiquitously expressed, heterotrimeric serine/threonine kinase whose short-term activation has multiple beneficial metabolic effects. Whether these translate into long-term benefits for obesity and its complications is unknown. Here, we observe that mice with chronic AMPK activation, resulting from mutation of the AMPK γ2 subunit, exhibit ghrelin signaling-dependent hyperphagia, obesity, and impaired pancreatic islet insulin secretion. Humans bearing the homologous mutation manifest a congruent phenotype. Our studies highlight that long-term AMPK activation throughout all tissues can have adverse metabolic consequences, with implications for pharmacological strategies seeking to chronically activate AMPK systemically to treat metabolic disease.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Andersson U., Filipsson K., Abbott C.R., Woods A., Smith K., Bloom S.R., Carling D., Small C.J. AMP-activated protein kinase plays a role in the control of food intake. J. Biol. Chem. 2004;279:12005–12008. - PubMed
-
- Bauer U.E., Briss P.A., Goodman R.A., Bowman B.A. Prevention of chronic disease in the 21st century: elimination of the leading preventable causes of premature death and disability in the USA. Lancet. 2014;384:45–52. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
