

Network Homeostasis and State Dynamics of Neocortical Sleep
- PMID: 27133462
- PMCID: PMC4873379
- DOI: 10.1016/j.neuron.2016.03.036
Network Homeostasis and State Dynamics of Neocortical Sleep
Abstract
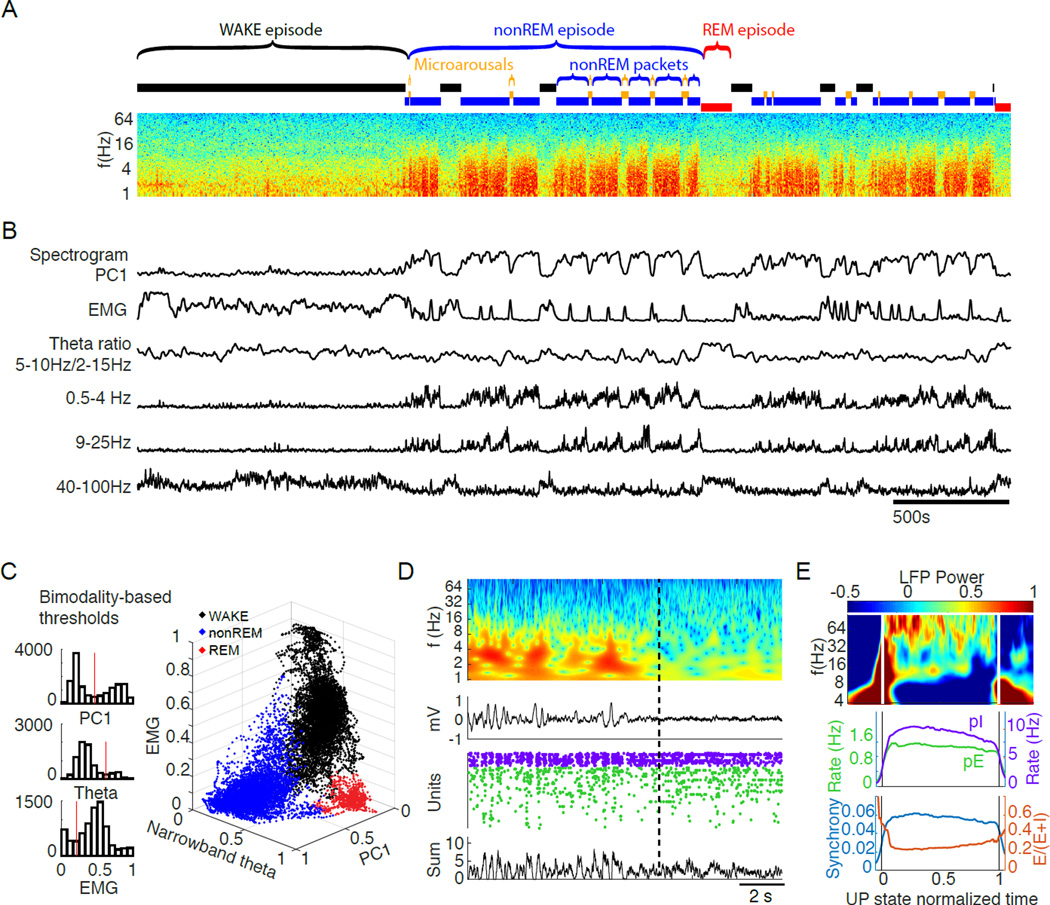
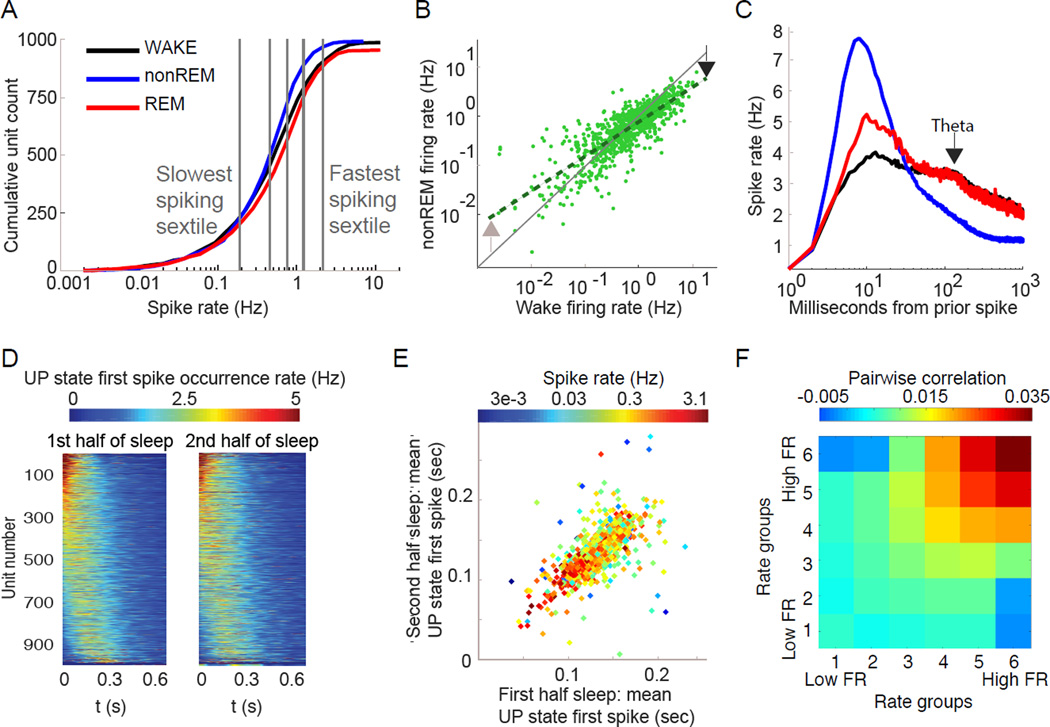
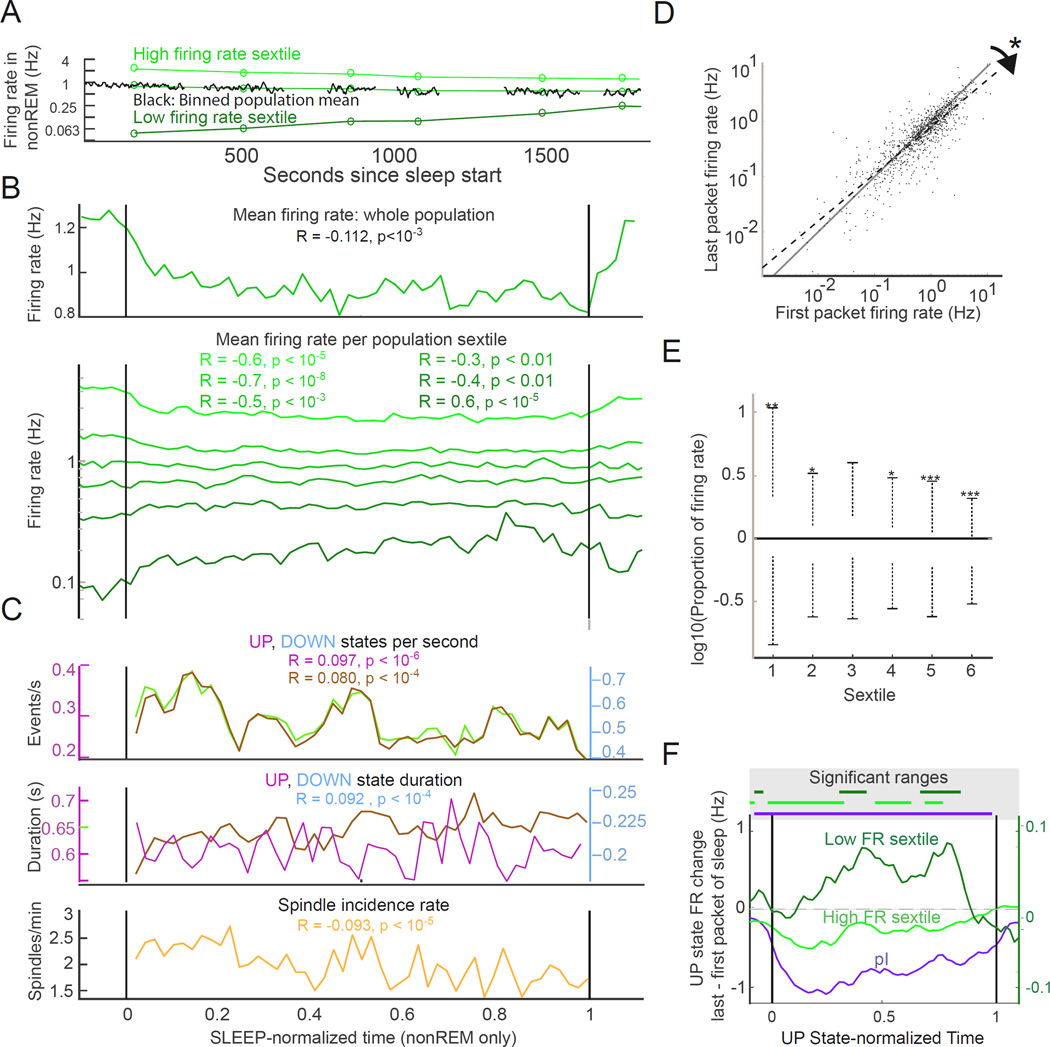
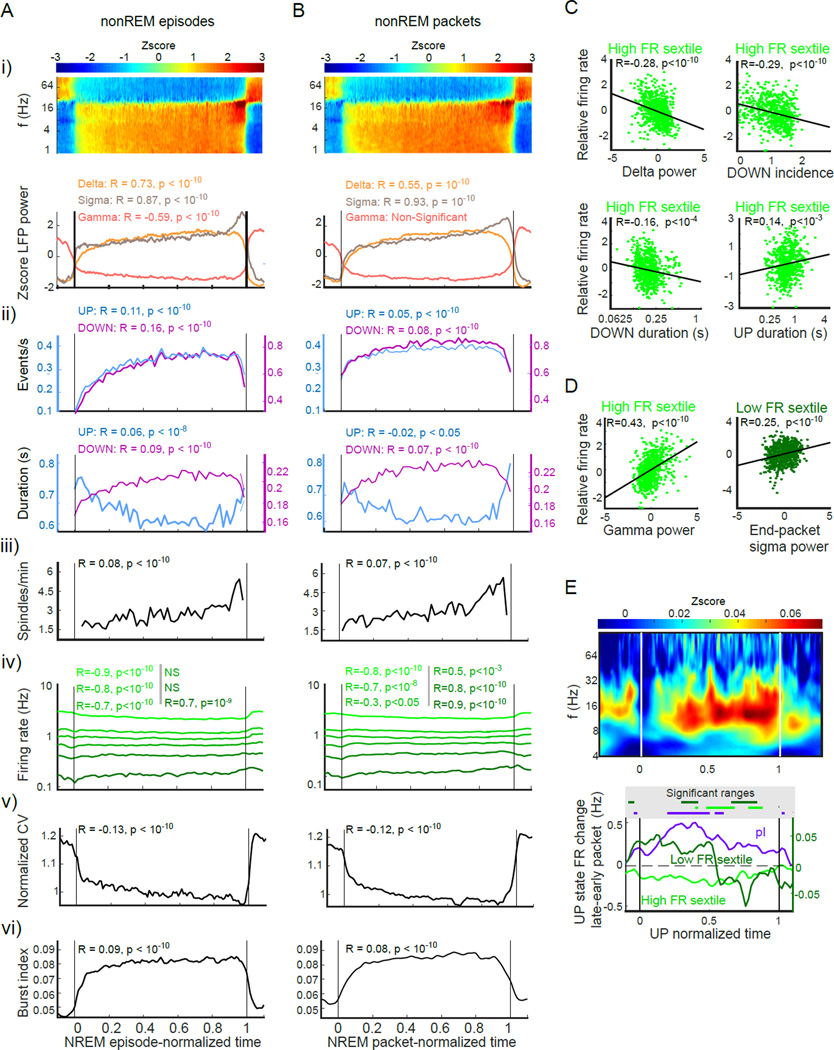
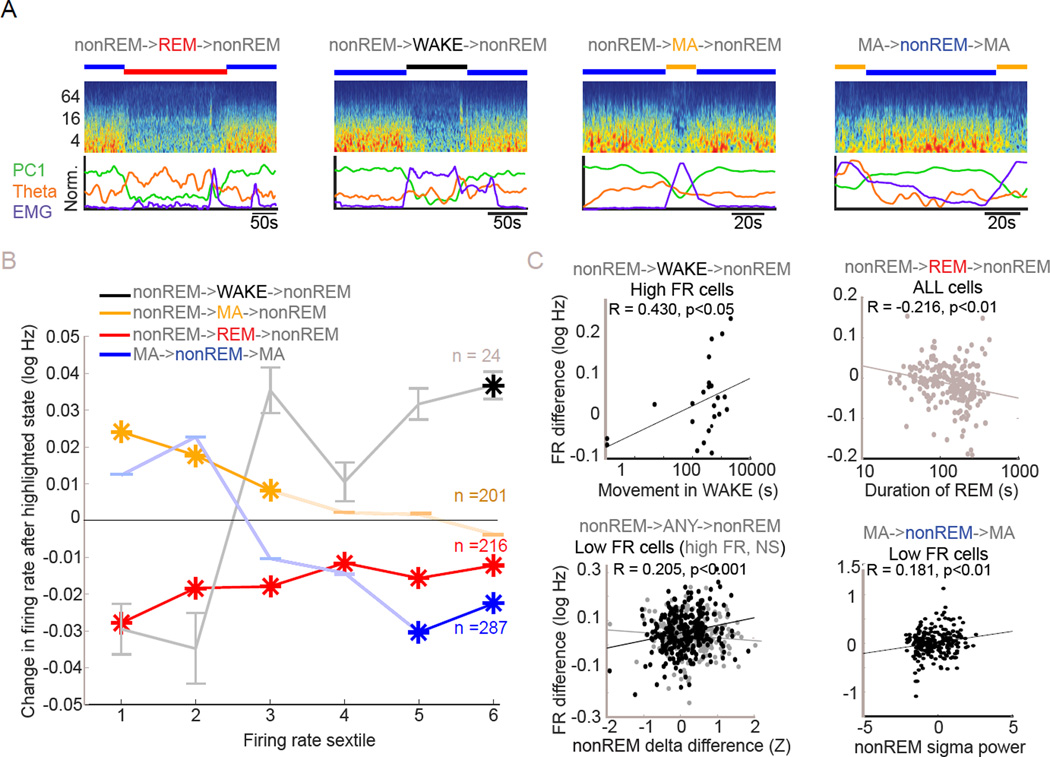
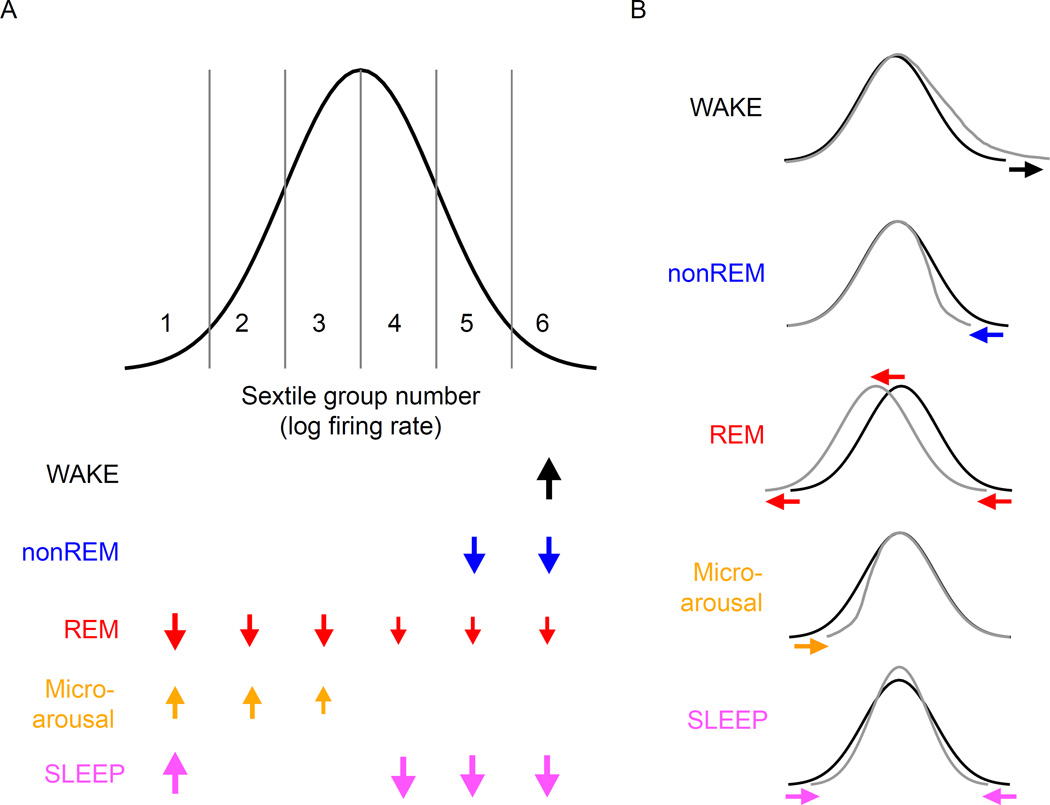
Sleep exerts many effects on mammalian forebrain networks, including homeostatic effects on both synaptic strengths and firing rates. We used large-scale recordings to examine the activity of neurons in the frontal cortex of rats and first observed that the distribution of pyramidal cell firing rates was wide and strongly skewed toward high firing rates. Moreover, neurons from different parts of that distribution were differentially modulated by sleep substates. Periods of nonREM sleep reduced the activity of high firing rate neurons and tended to upregulate firing of slow-firing neurons. By contrast, the effect of REM was to reduce firing rates across the entire rate spectrum. Microarousals, interspersed within nonREM epochs, increased firing rates of slow-firing neurons. The net result of sleep was to homogenize the firing rate distribution. These findings are at variance with current homeostatic models and provide a novel view of sleep in adjusting network excitability.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
