

Architecture of the mitochondrial calcium uniporter
- PMID: 27135929
- PMCID: PMC4874835
- DOI: 10.1038/nature17656
Architecture of the mitochondrial calcium uniporter
Abstract
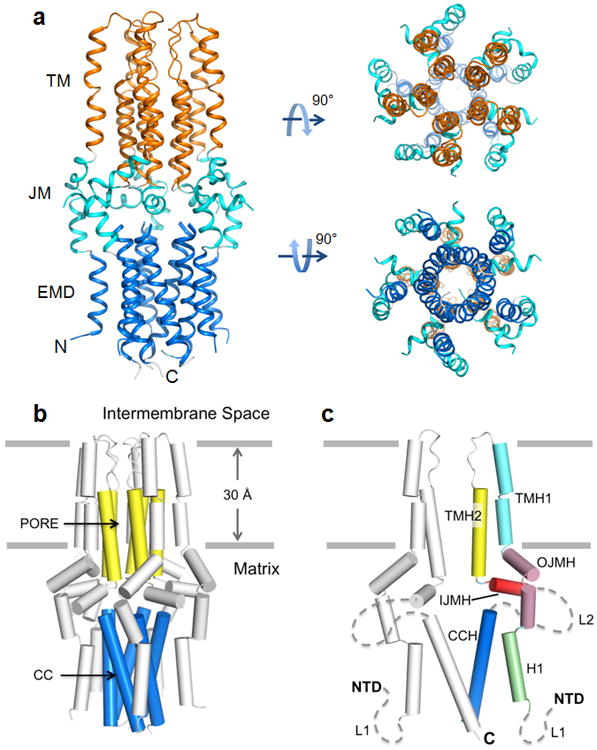
Mitochondria from many eukaryotic clades take up large amounts of calcium (Ca(2+)) via an inner membrane transporter called the uniporter. Transport by the uniporter is membrane potential dependent and sensitive to ruthenium red or its derivative Ru360 (ref. 1). Electrophysiological studies have shown that the uniporter is an ion channel with remarkably high conductance and selectivity. Ca(2+) entry into mitochondria is also known to activate the tricarboxylic acid cycle and seems to be crucial for matching the production of ATP in mitochondria with its cytosolic demand. Mitochondrial calcium uniporter (MCU) is the pore-forming and Ca(2+)-conducting subunit of the uniporter holocomplex, but its primary sequence does not resemble any calcium channel studied to date. Here we report the structure of the pore domain of MCU from Caenorhabditis elegans, determined using nuclear magnetic resonance (NMR) and electron microscopy (EM). MCU is a homo-oligomer in which the second transmembrane helix forms a hydrophilic pore across the membrane. The channel assembly represents a new solution of ion channel architecture, and is stabilized by a coiled-coil motif protruding into the mitochondrial matrix. The critical DXXE motif forms the pore entrance, which features two carboxylate rings; based on the ring dimensions and functional mutagenesis, these rings appear to form the selectivity filter. To our knowledge, this is one of the largest membrane protein structures characterized by NMR, and provides a structural blueprint for understanding the function of this channel.
Figures

 . The serine residue shown to be involved in Ru360 inhibition is indicated by *. Helical segments, as determined by NMR in this study, are indicated by cylinders and labeled as in the main text. The accession numbers for hMCU and cMCU are NM_138357.1 and NP_500892.1, respectively.
. The serine residue shown to be involved in Ru360 inhibition is indicated by *. Helical segments, as determined by NMR in this study, are indicated by cylinders and labeled as in the main text. The accession numbers for hMCU and cMCU are NM_138357.1 and NP_500892.1, respectively.










References
-
- Gunter TE, Pfeiffer DR. Mechanisms by which mitochondria transport calcium. The American journal of physiology. 1990;258:C755–786. - PubMed
-
- Kirichok Y, Krapivinsky G, Clapham DE. The mitochondrial calcium uniporter is a highly selective ion channel. Nature. 2004;427:360–364. - PubMed
-
- Denton RM, McCormack JG. The role of calcium in the regulation of mitochondrial metabolism. Biochem Soc Trans. 1980;8:266–268. - PubMed
Methods References
-
- Cong Y, Kovacs JA, Wriggers W. 2D fast rotational matching for image processing of biophysical data. Journal of structural biology. 2003;144:51–60. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
