

Characterization of MUDENG, a novel anti-apoptotic protein
- PMID: 27136675
- PMCID: PMC4945747
- DOI: 10.1038/oncsis.2016.30
Characterization of MUDENG, a novel anti-apoptotic protein
Abstract
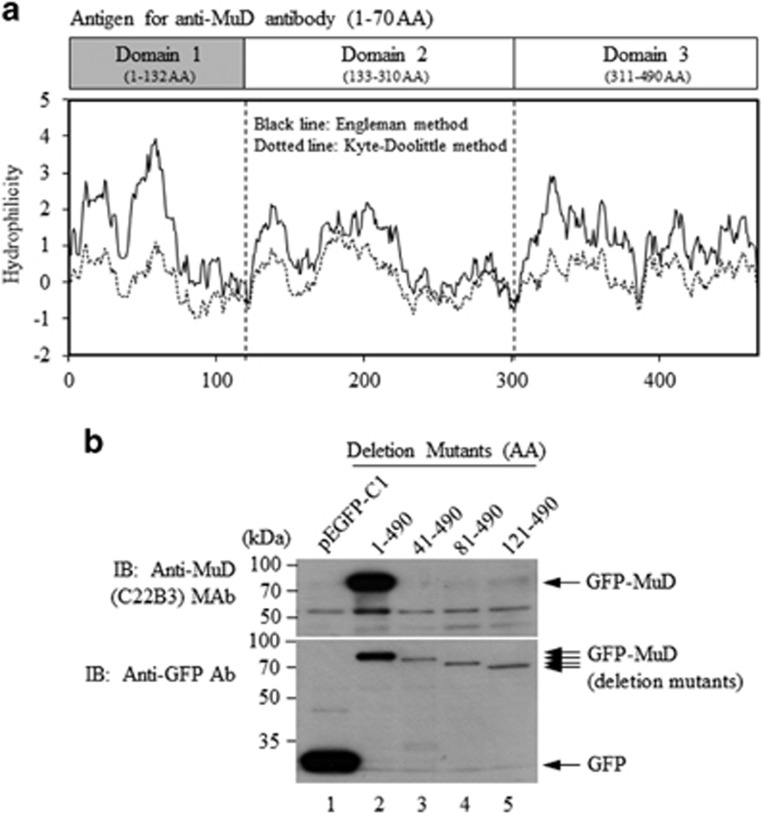
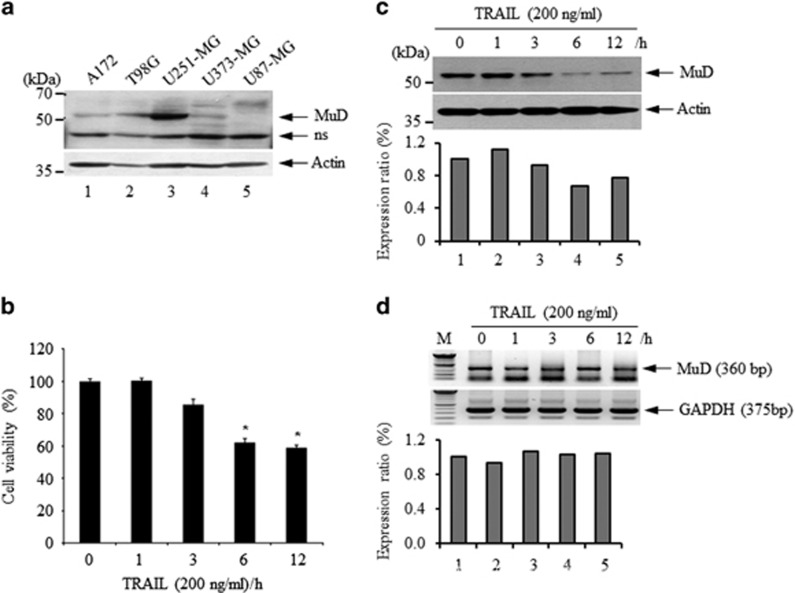
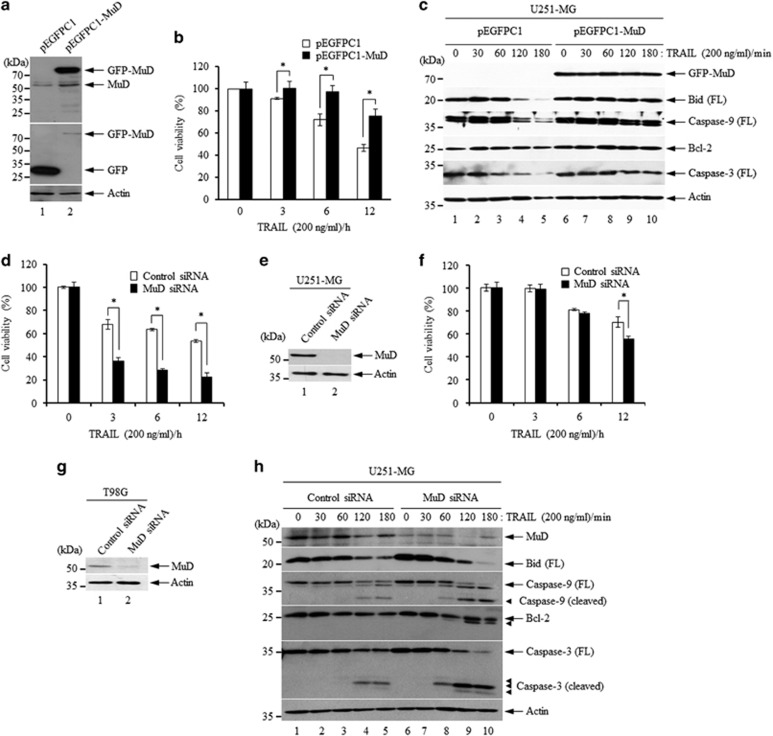
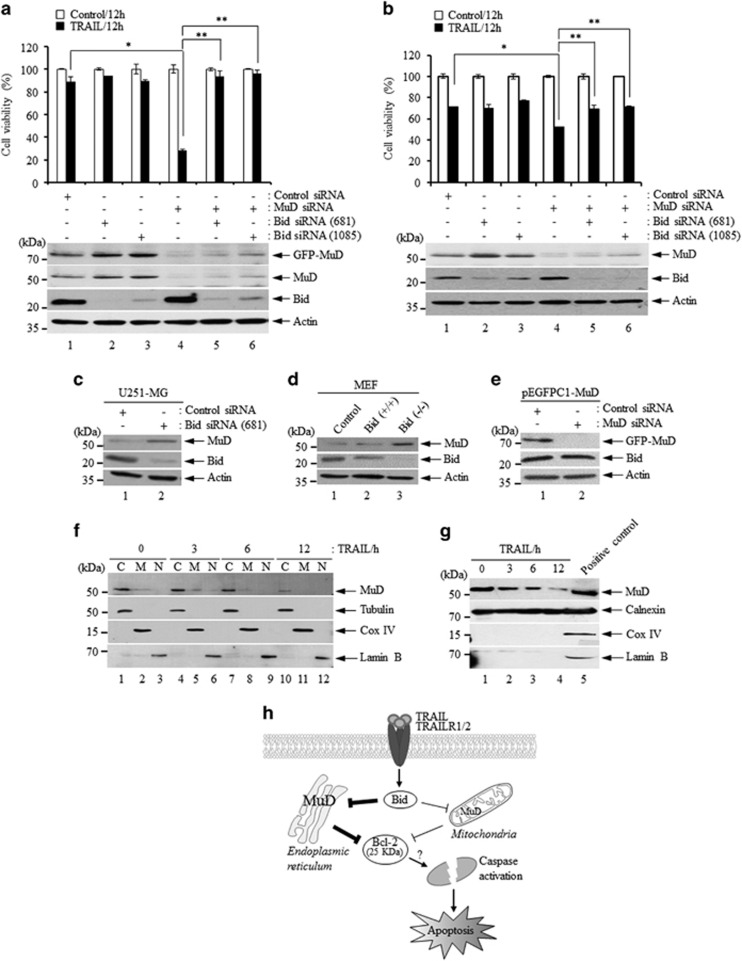
MUDENG (Mu-2-related death-inducing gene, MuD) is revealed to be involved in cell death signaling. Astrocytes, the major glial cell type in the central nervous system, are a source of brain tumors. In this study, we examined MuD expression and function in human astroglioma cells. Stimulation of U251-MG cells with tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) resulted in a 40% decrease in cell viability and a 33% decrease in MuD protein levels, although not in MuD mRNA levels. To study the functional relevance of MuD expression, stable transfectants expressing high levels of MuD were generated. Stimulation of these transfectants with TRAIL resulted in enhanced cell survival (77% for stable and 46% for control transfectants). Depletion of MuD led to a marked reduction upon TRAIL stimulation in cell viability (22% in MuD-depleted cells and 54% in control cells). In addition, we observed that MuD depletion increased the susceptibility of the cells to TRAIL by enhancing the cleavage of caspase-3/-9 and BH3-interacting domain death agonist (Bid). A unique 25-kDa fragment of B-cell lymphoma 2 (Bcl-2) lacking BH4 was observed 60-180 min post TRAIL treatment in MuD-depleted cells, suggesting that Bcl-2 is converted from its anti-apoptotic form to the truncated pro-apoptotic form. Importantly, the TRAIL-mediated decrease in cell viability in MuD-depleted cells was abrogated upon Bid depletion, indicating that the role of MuD in apoptotic signaling takes place at the Bid and Bcl-2 junction. MuD localizes predominantly in the endoplasmic reticulum and partly in the mitochondria and its amounts are reduced 6 h post TRAIL stimulation, presumably via caspase-3-mediated MuD cleavage. Collectively, these results suggest that MuD, a novel signaling protein, not only possesses an anti-apoptotic function but may also constitute an important target for the design of ideal candidates for combinatorial treatment strategies for glioma cells.
Figures
Similar articles
-
The graviola impact on human astroglioma cells: functional significance of MUDENG.RSC Adv. 2019 Mar 18;9(16):8935-8942. doi: 10.1039/c8ra10039j. eCollection 2019 Mar 15. RSC Adv. 2019. PMID: 35517667 Free PMC article.
-
Silver nanoparticle-induced hormesis of astroglioma cells: A Mu-2-related death-inducing protein-orchestrated modus operandi.Int J Biol Macromol. 2018 Oct 1;117:1147-1156. doi: 10.1016/j.ijbiomac.2018.05.234. Epub 2018 Jun 2. Int J Biol Macromol. 2018. PMID: 29870812
-
A caspase-8-independent component in TRAIL/Apo-2L-induced cell death in human rhabdomyosarcoma cells.Cell Death Differ. 2003 Jun;10(6):729-39. doi: 10.1038/sj.cdd.4401232. Cell Death Differ. 2003. PMID: 12761581
-
The MUDENG Augmentation: A Genesis in Anti-Cancer Therapy?Int J Mol Sci. 2020 Aug 4;21(15):5583. doi: 10.3390/ijms21155583. Int J Mol Sci. 2020. PMID: 32759789 Free PMC article. Review.
-
Down-regulation of intracellular anti-apoptotic proteins, particularly c-FLIP by therapeutic agents; the novel view to overcome resistance to TRAIL.J Cell Physiol. 2018 Oct;233(10):6470-6485. doi: 10.1002/jcp.26585. Epub 2018 May 9. J Cell Physiol. 2018. PMID: 29741767 Review.
Cited by
-
Quercetin-3-Glucoside Extracted from Apple Pomace Induces Cell Cycle Arrest and Apoptosis by Increasing Intracellular ROS Levels.Int J Mol Sci. 2021 Oct 4;22(19):10749. doi: 10.3390/ijms221910749. Int J Mol Sci. 2021. PMID: 34639090 Free PMC article.
-
Gintonin-Enriched Panax ginseng Extract Fraction Sensitizes Renal Carcinoma Cells to TRAIL-Induced Apoptosis through DR4/5 Upregulation.Curr Issues Mol Biol. 2024 Sep 27;46(10):10880-10895. doi: 10.3390/cimb46100646. Curr Issues Mol Biol. 2024. PMID: 39451526 Free PMC article.
-
Role of adaptin protein complexes in intracellular trafficking and their impact on diseases.Bioengineered. 2021 Dec;12(1):8259-8278. doi: 10.1080/21655979.2021.1982846. Bioengineered. 2021. PMID: 34565296 Free PMC article.
-
Generation of single-chain Fv antibody fragments against Mu-2-related death-inducing gene in Escherichia coli.Mol Biol Rep. 2019 Aug;46(4):4027-4037. doi: 10.1007/s11033-019-04852-1. Epub 2019 May 9. Mol Biol Rep. 2019. PMID: 31073914
-
Cinnamaldehyde-Rich Cinnamon Extract Induces Cell Death in Colon Cancer Cell Lines HCT 116 and HT-29.Int J Mol Sci. 2023 May 3;24(9):8191. doi: 10.3390/ijms24098191. Int J Mol Sci. 2023. PMID: 37175897 Free PMC article.
References
-
- Huse JT, Holland EC. Targeting brain cancer: advances in the molecular pathology of malignant glioma and medulloblastoma. Nat Rev Cancer 2010; 10: 319–331. - PubMed
-
- Kuijlen JM, Bremer E, Mooij JJ, den Dunnen WF, Helfrich W. Review: on TRAIL for malignant glioma therapy? Neuropathol Appl Neurobiol 2010; 36: 168–182. - PubMed
-
- Stewart LA. Chemotherapy in adult high-grade glioma: a systematic review and meta-analysis of individual patient data from 12 randomised trials. Lancet 2002; 359: 1011–1018. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
