

A novel lipid transfer protein from the pea Pisum sativum: isolation, recombinant expression, solution structure, antifungal activity, lipid binding, and allergenic properties
- PMID: 27137920
- PMCID: PMC4852415
- DOI: 10.1186/s12870-016-0792-6
A novel lipid transfer protein from the pea Pisum sativum: isolation, recombinant expression, solution structure, antifungal activity, lipid binding, and allergenic properties
Abstract
Background: Plant lipid transfer proteins (LTPs) assemble a family of small (7-9 kDa) ubiquitous cationic proteins with an ability to bind and transport lipids as well as participate in various physiological processes including defense against phytopathogens. They also form one of the most clinically relevant classes of plant allergens. Nothing is known to date about correlation between lipid-binding and IgE-binding properties of LTPs. The garden pea Pisum sativum is widely consumed crop and important allergenic specie of the legume family. This work is aimed at isolation of a novel LTP from pea seeds and characterization of its structural, functional, and allergenic properties.
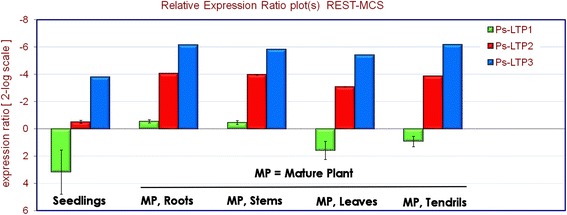
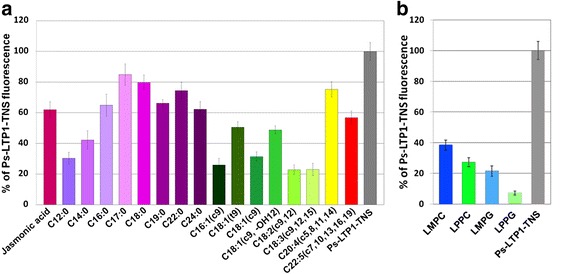
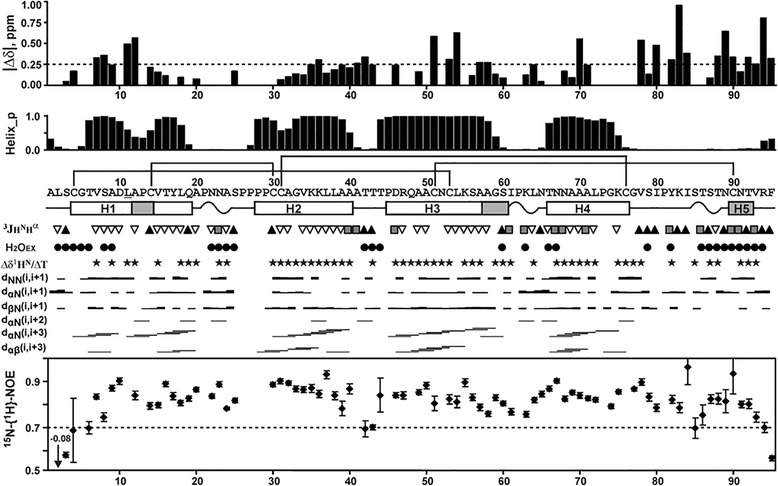
Results: Three novel lipid transfer proteins, designated as Ps-LTP1-3, were found in the garden pea Pisum sativum, their cDNA sequences were determined, and mRNA expression levels of all the three proteins were measured at different pea organs. Ps-LTP1 was isolated for the first time from the pea seeds, and its complete amino acid sequence was determined. The protein exhibits antifungal activity and is a membrane-active compound that causes a leakage from artificial liposomes. The protein binds various lipids including bioactive jasmonic acid. Spatial structure of the recombinant uniformly (13)C,(15)N-labelled Ps-LTP1 was solved by heteronuclear NMR spectroscopy. In solution the unliganded protein represents the mixture of two conformers (relative populations ~ 85:15) which are interconnected by exchange process with characteristic time ~ 100 ms. Hydrophobic residues of major conformer form a relatively large internal tunnel-like lipid-binding cavity (van der Waals volume comes up to ~1000 Å(3)). The minor conformer probably corresponds to the protein with the partially collapsed internal cavity.
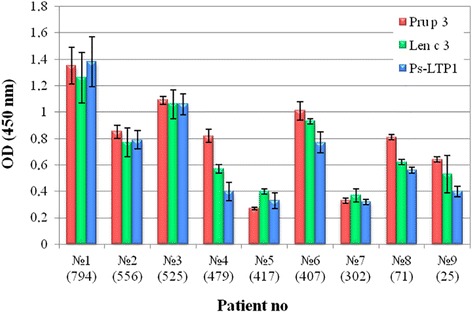
Conclusions: For the first time conformational heterogeneity in solution was shown for an unliganded plant lipid transfer protein. Heat denaturation profile and simulated gastrointestinal digestion assay showed that Ps-LTP1 displayed a high thermal and digestive proteolytic resistance proper for food allergens. The reported structural and immunological findings seem to describe Ps-LTP1 as potential cross-reactive allergen in LTP-sensitized patients, mostly Pru p 3(+) ones. Similarly to allergenic LTPs the potential IgE-binding epitope of Ps-LTP1 is located near the proposed entrance into internal cavity and could be involved in lipid-binding.
Keywords: Allergen; Antimicrobial activity; Differential gene expression; Garden pea; Heteronuclear NMR spectroscopy; Lipid binding; Lipid transfer protein; Pisum sativum; Recombinant expression; Spatial structure.
Figures



References
-
- Salcedo G, Sánchez-Monge R, Barber D, Díaz-Perales A. Plant non-specific lipid transfer proteins: an interface between plant defence and human allergy. Biochim Biophys Acta. 1771;2007:781–91. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
