

Experimental evolution reveals that high relatedness protects multicellular cooperation from cheaters
- PMID: 27139112
- PMCID: PMC4857390
- DOI: 10.1038/ncomms11435
Experimental evolution reveals that high relatedness protects multicellular cooperation from cheaters
Abstract
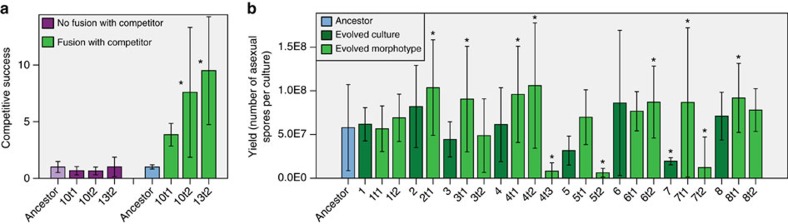
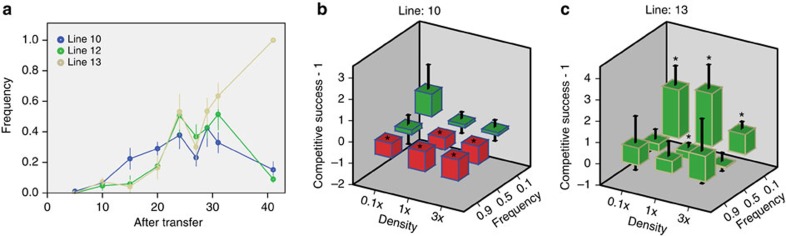
In multicellular organisms, there is a potential risk that cheating mutants gain access to the germline. Development from a single-celled zygote resets relatedness among cells to its maximum value each generation, which should accomplish segregation of cheating mutants from non-cheaters and thereby protect multicellular cooperation. Here we provide the crucial direct comparison between high- and low-relatedness conditions to test this hypothesis. We allow two variants of the fungus Neurospora crassa to evolve, one with and one without the ability to form chimeras with other individuals, thus generating two relatedness levels. While multicellular cooperation remains high in the high-relatedness lines, it significantly decreases in all replicate low-relatedness lines, resulting in an average threefold decrease in spore yield. This reduction is caused by cheating mutants with reduced investment in somatic functions, but increased competitive success when fusing with non-cheaters. Our experiments demonstrate that high genetic relatedness is crucial to sustain multicellular cooperation.
Figures




Similar articles
-
High relatedness maintains multicellular cooperation in a social amoeba by controlling cheater mutants.Proc Natl Acad Sci U S A. 2007 May 22;104(21):8913-7. doi: 10.1073/pnas.0702723104. Epub 2007 May 11. Proc Natl Acad Sci U S A. 2007. PMID: 17496139 Free PMC article.
-
Does high relatedness promote cheater-free multicellularity in synthetic lifecycles?J Evol Biol. 2017 May;30(5):985-993. doi: 10.1111/jeb.13067. Epub 2017 Apr 7. J Evol Biol. 2017. PMID: 28294448
-
Concurrent coevolution of intra-organismal cheaters and resisters.J Evol Biol. 2015 Apr;28(4):756-65. doi: 10.1111/jeb.12618. Epub 2015 Mar 28. J Evol Biol. 2015. PMID: 25772340
-
Evolution of cooperation and control of cheating in a social microbe.Proc Natl Acad Sci U S A. 2011 Jun 28;108 Suppl 2(Suppl 2):10855-62. doi: 10.1073/pnas.1102451108. Epub 2011 Jun 20. Proc Natl Acad Sci U S A. 2011. PMID: 21690338 Free PMC article. Review.
-
Genetic control of asexual sporulation in filamentous fungi.Curr Opin Microbiol. 2012 Dec;15(6):669-77. doi: 10.1016/j.mib.2012.09.006. Epub 2012 Oct 22. Curr Opin Microbiol. 2012. PMID: 23092920 Review.
Cited by
-
Multispecies interactions shape the transition to multicellularity.Proc Biol Sci. 2023 Sep 27;290(2007):20231055. doi: 10.1098/rspb.2023.1055. Epub 2023 Sep 20. Proc Biol Sci. 2023. PMID: 37727086 Free PMC article.
-
Identification of a common secondary mutation in the Neurospora crassa knockout collection conferring a cell fusion-defective phenotype.Microbiol Spectr. 2023 Aug 25;11(5):e0208723. doi: 10.1128/spectrum.02087-23. Online ahead of print. Microbiol Spectr. 2023. PMID: 37623742 Free PMC article.
-
Ten recent insights for our understanding of cooperation.Nat Ecol Evol. 2021 Apr;5(4):419-430. doi: 10.1038/s41559-020-01384-x. Epub 2021 Jan 28. Nat Ecol Evol. 2021. PMID: 33510431 Free PMC article. Review.
-
Molecular Mechanisms Regulating Cell Fusion and Heterokaryon Formation in Filamentous Fungi.Microbiol Spectr. 2017 Mar;5(2):10.1128/microbiolspec.funk-0015-2016. doi: 10.1128/microbiolspec.FUNK-0015-2016. Microbiol Spectr. 2017. PMID: 28256191 Free PMC article. Review.
-
Adaptation to public goods cheats in Pseudomonas aeruginosa.Proc Biol Sci. 2017 Jul 26;284(1859):20171089. doi: 10.1098/rspb.2017.1089. Proc Biol Sci. 2017. PMID: 28747481 Free PMC article.
References
-
- Buss L. The Evolution of Individuality Princeton University Press (1987).
-
- Maynard Smith J. & Szathmary E. The Major Transitions in Evolution Oxford University Press (1995).
-
- Bonner J. On the origin of differentiation. J. Biosci. 28, 523–528 (2003). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
