

Characterization of primary human hepatocyte spheroids as a model system for drug-induced liver injury, liver function and disease
- PMID: 27143246
- PMCID: PMC4855186
- DOI: 10.1038/srep25187
Characterization of primary human hepatocyte spheroids as a model system for drug-induced liver injury, liver function and disease
Abstract
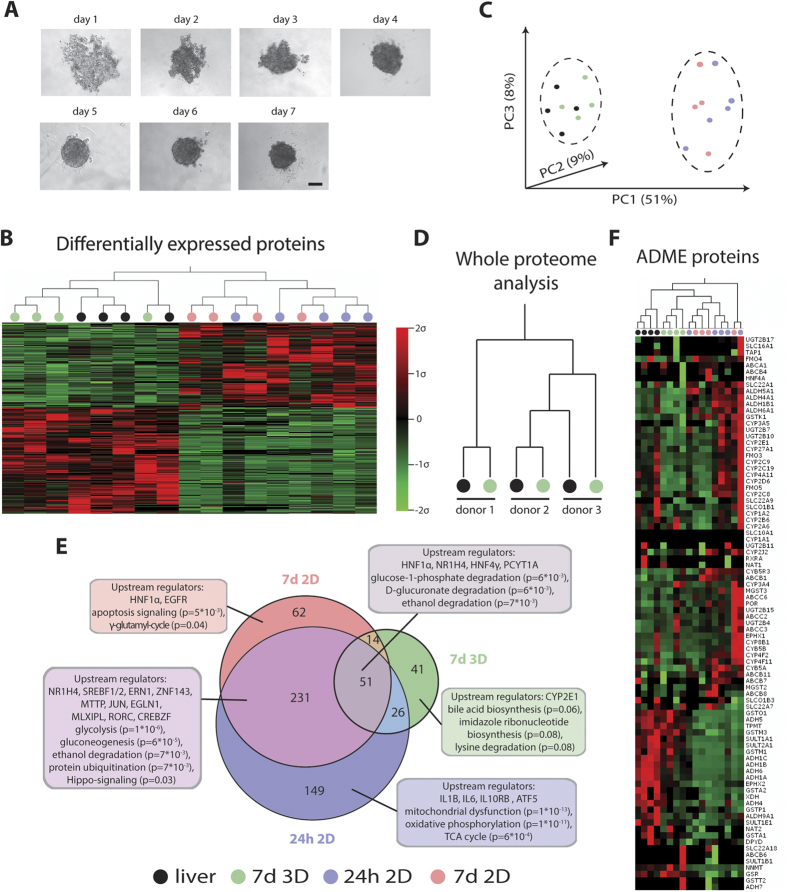
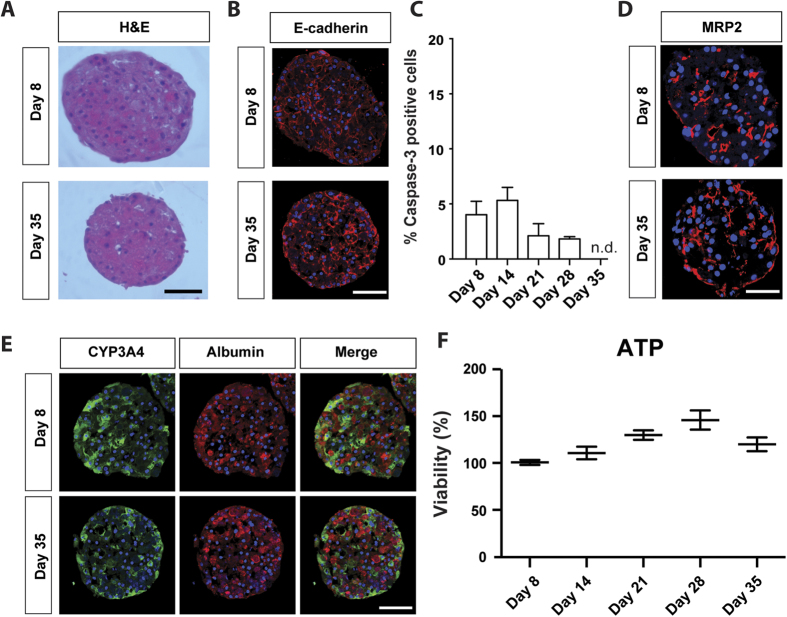
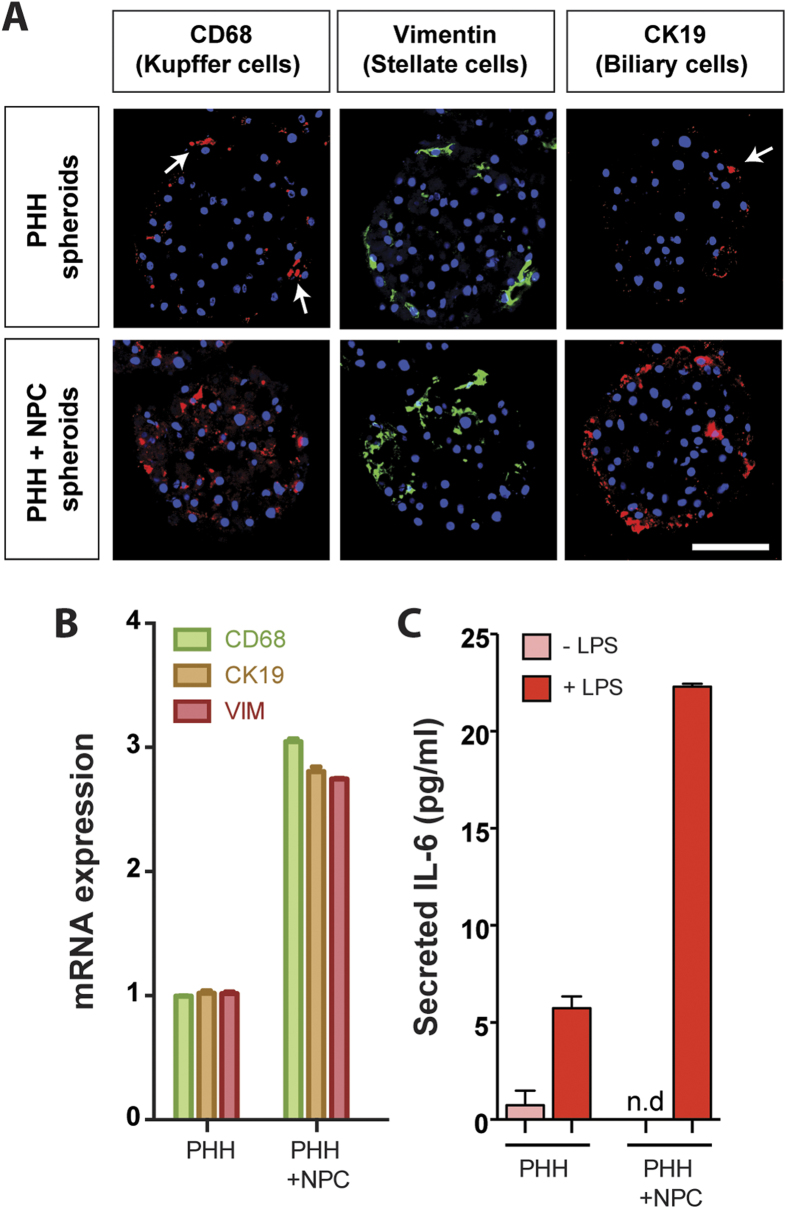
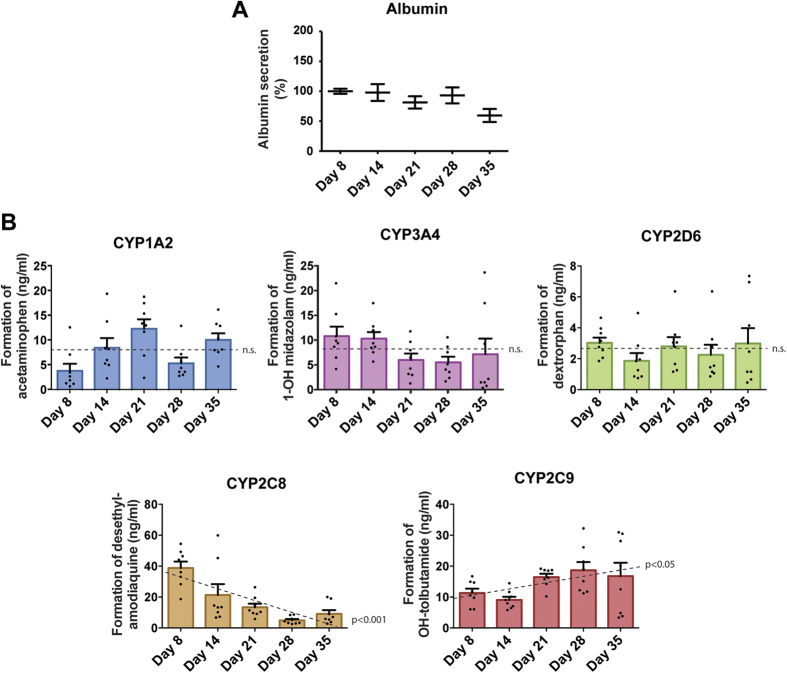
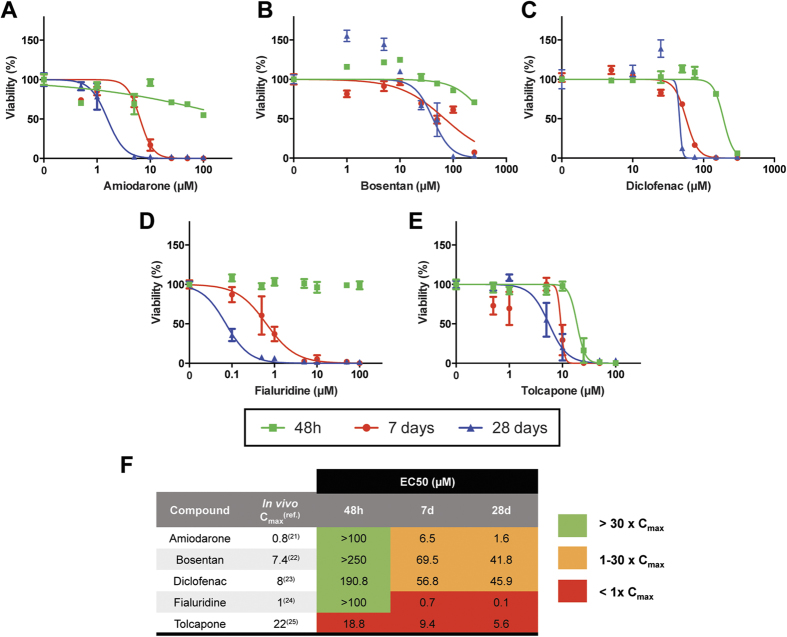
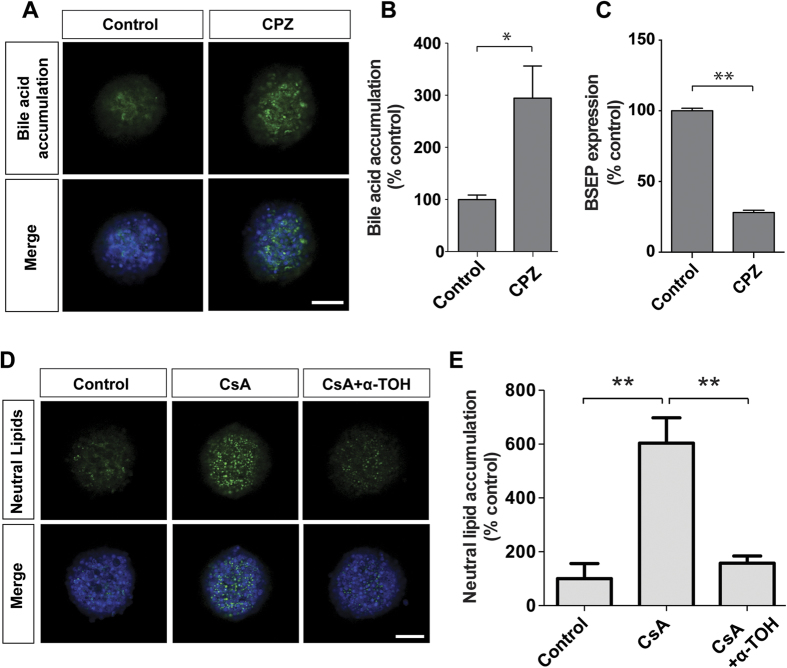
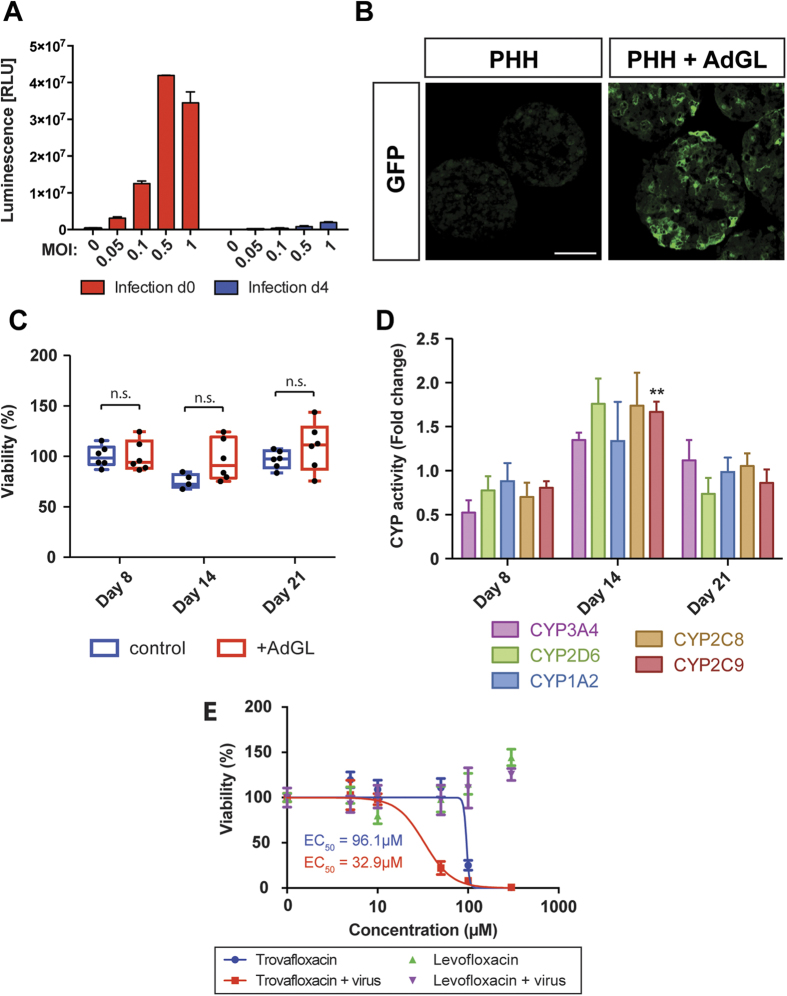
Liver biology and function, drug-induced liver injury (DILI) and liver diseases are difficult to study using current in vitro models such as primary human hepatocyte (PHH) monolayer cultures, as their rapid de-differentiation restricts their usefulness substantially. Thus, we have developed and extensively characterized an easily scalable 3D PHH spheroid system in chemically-defined, serum-free conditions. Using whole proteome analyses, we found that PHH spheroids cultured this way were similar to the liver in vivo and even retained their inter-individual variability. Furthermore, PHH spheroids remained phenotypically stable and retained morphology, viability, and hepatocyte-specific functions for culture periods of at least 5 weeks. We show that under chronic exposure, the sensitivity of the hepatocytes drastically increased and toxicity of a set of hepatotoxins was detected at clinically relevant concentrations. An interesting example was the chronic toxicity of fialuridine for which hepatotoxicity was mimicked after repeated-dosing in the PHH spheroid model, not possible to detect using previous in vitro systems. Additionally, we provide proof-of-principle that PHH spheroids can reflect liver pathologies such as cholestasis, steatosis and viral hepatitis. Combined, our results demonstrate that the PHH spheroid system presented here constitutes a versatile and promising in vitro system to study liver function, liver diseases, drug targets and long-term DILI.
Figures
References
-
- Hewitt N. J. et al. Primary Hepatocytes: Current Understanding of the Regulation of Metabolic Enzymes and Transporter Proteins, and Pharmaceutical Practice for the Use of Hepatocytes in Metabolism, Enzyme Induction, Transporter, Clearance, and Hepatotoxicity Studies. Drug Metab. Rev. 39, 159–234, 10.1080/03602530601093489 (2015). - DOI - PubMed
-
- Elaut G. et al. Molecular mechanisms underlying the dedifferentiation process of isolated hepatocytes and their cultures. Curr. Drug Metab. 7, 629–660 (2006). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
