

An FGF3-BMP Signaling Axis Regulates Caudal Neural Tube Closure, Neural Crest Specification and Anterior-Posterior Axis Extension
- PMID: 27144312
- PMCID: PMC4856314
- DOI: 10.1371/journal.pgen.1006018
An FGF3-BMP Signaling Axis Regulates Caudal Neural Tube Closure, Neural Crest Specification and Anterior-Posterior Axis Extension
Abstract
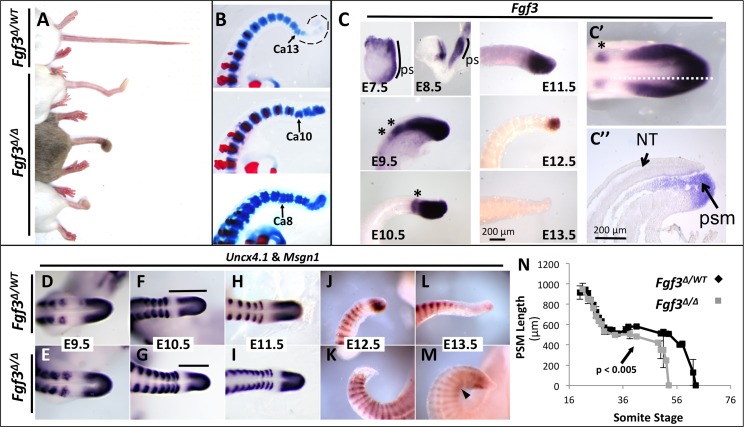
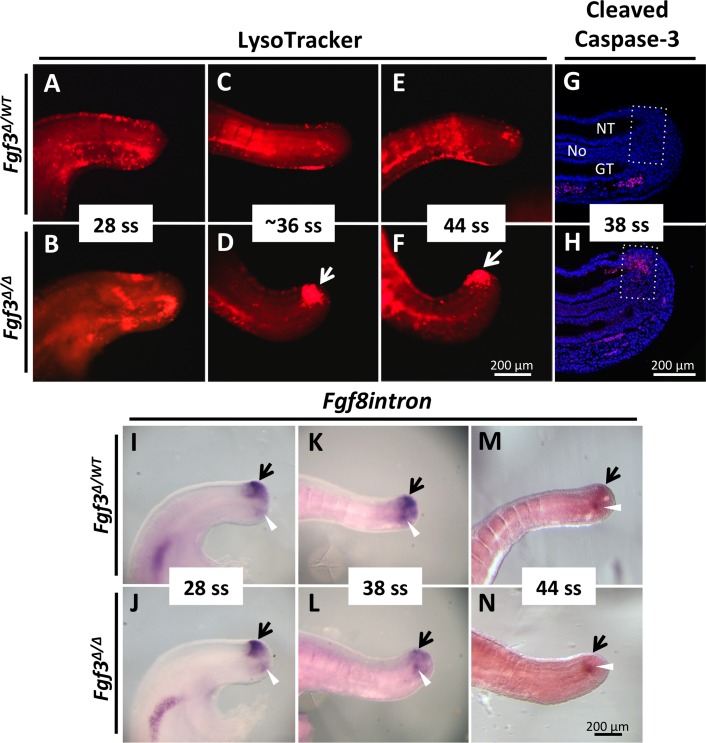
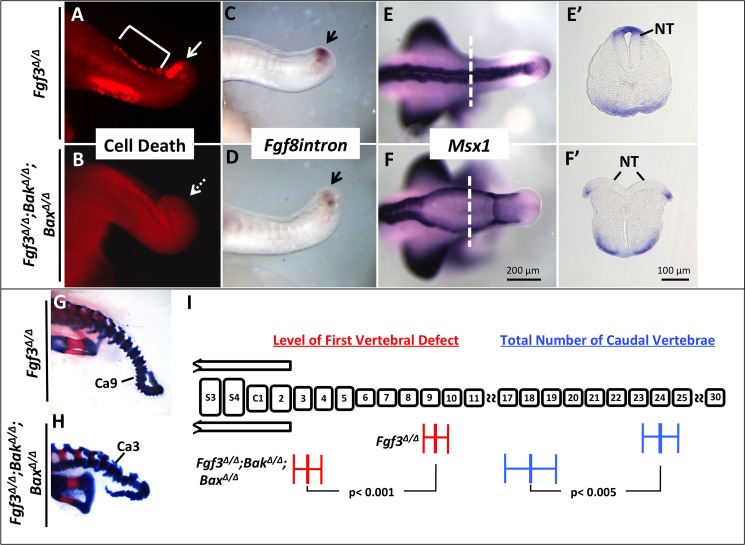
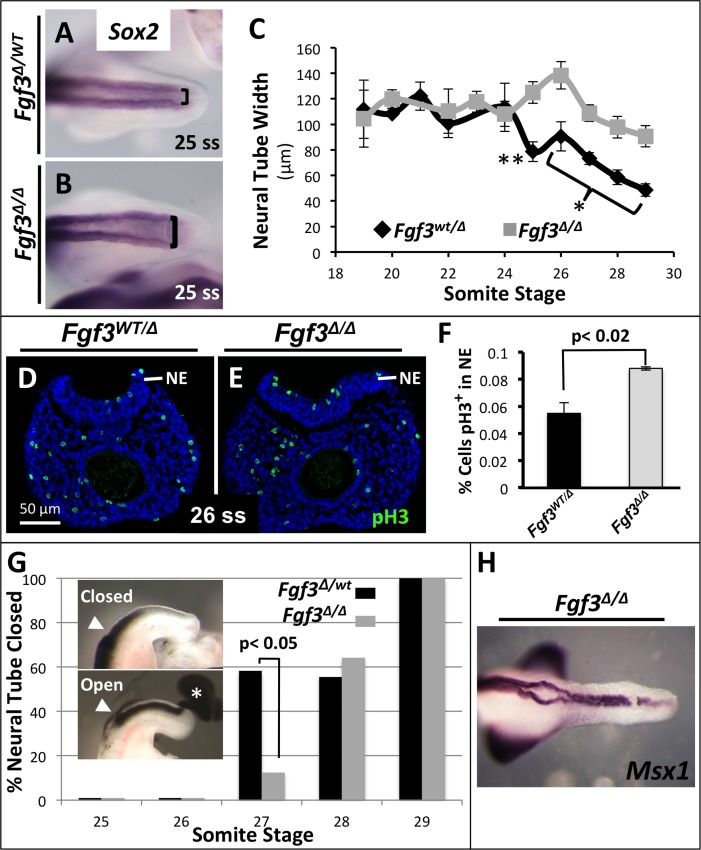
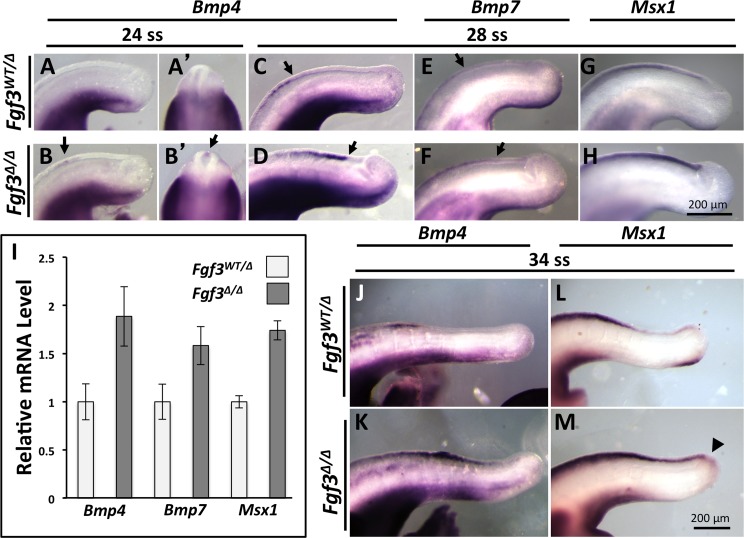
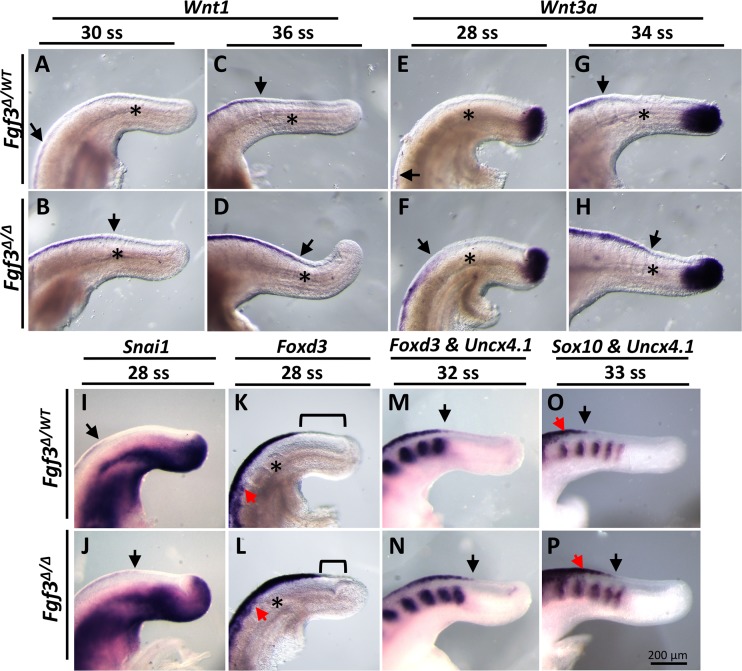
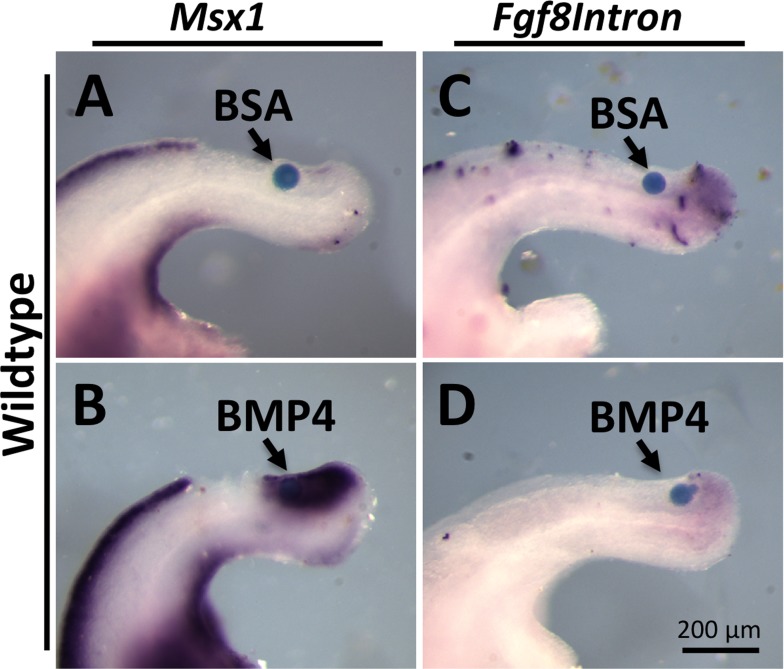
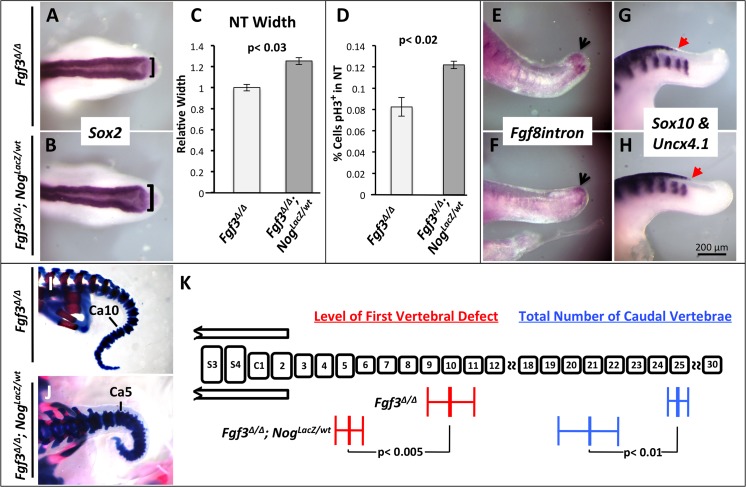
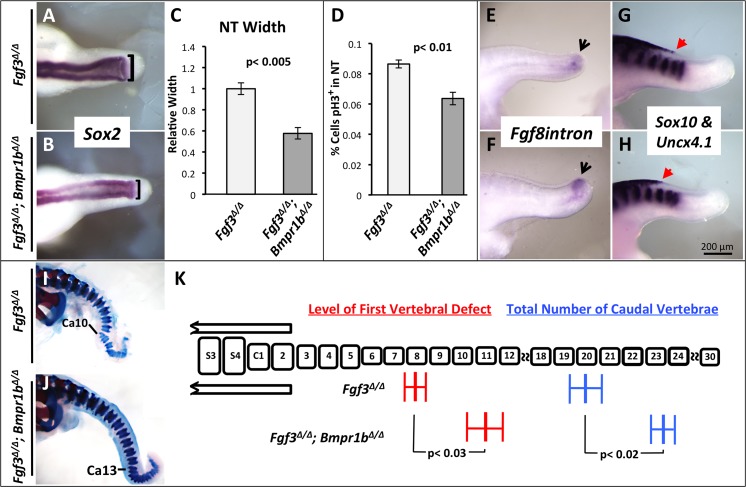
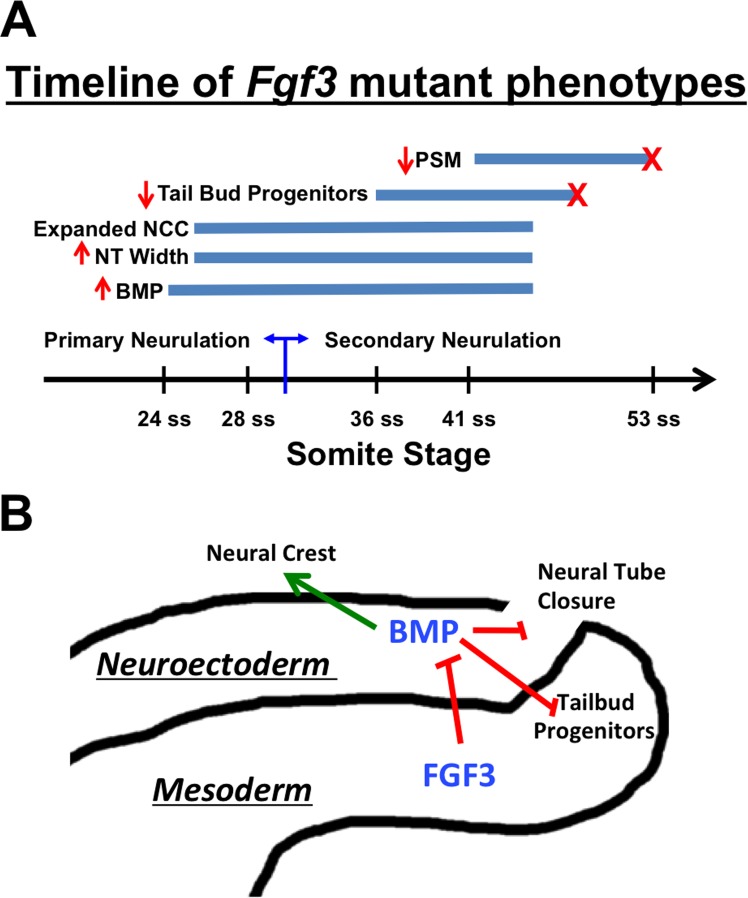
During vertebrate axis extension, adjacent tissue layers undergo profound morphological changes: within the neuroepithelium, neural tube closure and neural crest formation are occurring, while within the paraxial mesoderm somites are segmenting from the presomitic mesoderm (PSM). Little is known about the signals between these tissues that regulate their coordinated morphogenesis. Here, we analyze the posterior axis truncation of mouse Fgf3 null homozygotes and demonstrate that the earliest role of PSM-derived FGF3 is to regulate BMP signals in the adjacent neuroepithelium. FGF3 loss causes elevated BMP signals leading to increased neuroepithelium proliferation, delay in neural tube closure and premature neural crest specification. We demonstrate that elevated BMP4 depletes PSM progenitors in vitro, phenocopying the Fgf3 mutant, suggesting that excessive BMP signals cause the Fgf3 axis defect. To test this in vivo we increased BMP signaling in Fgf3 mutants by removing one copy of Noggin, which encodes a BMP antagonist. In such mutants, all parameters of the Fgf3 phenotype were exacerbated: neural tube closure delay, premature neural crest specification, and premature axis termination. Conversely, genetically decreasing BMP signaling in Fgf3 mutants, via loss of BMP receptor activity, alleviates morphological defects. Aberrant apoptosis is observed in the Fgf3 mutant tailbud. However, we demonstrate that cell death does not cause the Fgf3 phenotype: blocking apoptosis via deletion of pro-apoptotic genes surprisingly increases all Fgf3 defects including causing spina bifida. We demonstrate that this counterintuitive consequence of blocking apoptosis is caused by the increased survival of BMP-producing cells in the neuroepithelium. Thus, we show that FGF3 in the caudal vertebrate embryo regulates BMP signaling in the neuroepithelium, which in turn regulates neural tube closure, neural crest specification and axis termination. Uncovering this FGF3-BMP signaling axis is a major advance toward understanding how these tissue layers interact during axis extension with important implications in human disease.
Conflict of interest statement
The authors have declared that no competing interests exist
Figures
References
-
- Zohn IE (2012) Mouse as a model for multifactorial inheritance of neural tube defects. Birth defects research Part C, Embryo today: reviews 96: 193–205. - PubMed
-
- Harris MJ, Juriloff DM (2007) Mouse mutants with neural tube closure defects and their role in understanding human neural tube defects. Birth defects research Part A, Clinical and molecular teratology 79: 187–210. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
