

Imbalanced Hemolymph Lipid Levels Affect Feeding Motivation in the Two-Spotted Cricket, Gryllus bimaculatus
- PMID: 27144650
- PMCID: PMC4856397
- DOI: 10.1371/journal.pone.0154841
Imbalanced Hemolymph Lipid Levels Affect Feeding Motivation in the Two-Spotted Cricket, Gryllus bimaculatus
Abstract
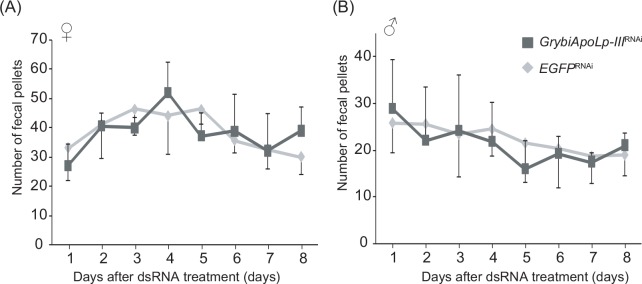
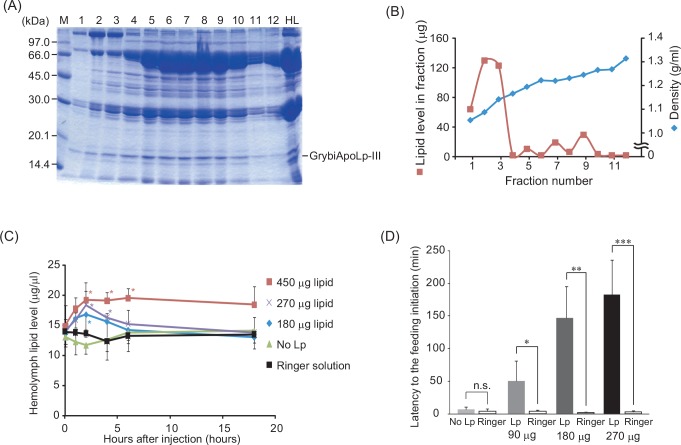
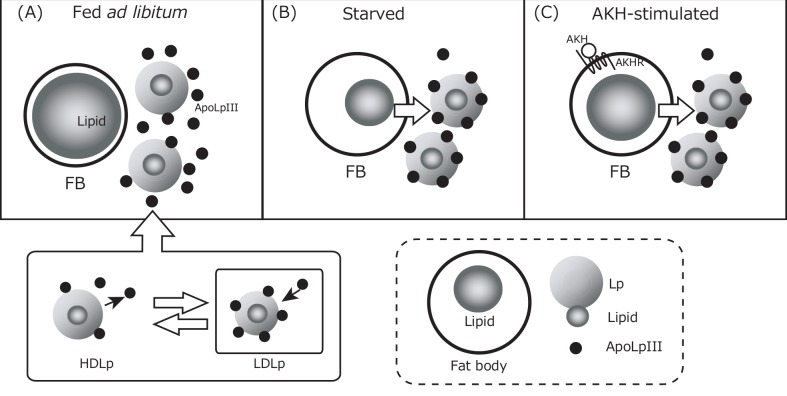
Insect feeding behavior is regulated by many intrinsic factors, including hemolymph nutrient levels. Adipokinetic hormone (AKH) is a peptide factor that modulates hemolymph nutrient levels and regulates the nutritional state of insects by triggering the transfer of lipids into the hemolymph. We recently demonstrated that RNA interference (RNAi)-mediated knockdown of the AKH receptor (AKHR) reduces hemolymph lipid levels, causing an increase in the feeding frequency of the two-spotted cricket, Gryllus bimaculatus. This result indicated that reduced hemolymph lipid levels might motivate crickets to feed. In the present study, to elucidate whether hemolymph lipid levels contribute to insect feeding behavior, we attempted to manipulate hemolymph lipid levels via the lipophorin (Lp)-mediated lipid transferring system in G. bimaculatus. Of the constituent proteins in Lp, we focused on apolipophorin-III (GrybiApoLp-III) because of its possible role in facilitating lipid mobilization. First, we used RNAi to reduce the expression of GrybiApoLp-III. RNAi-mediated knockdown of GrybiApoLp-III had little effect on basal hemolymph lipid levels and the amount of food intake. In addition, hemolymph lipid levels remained static even after injecting AKH into GrybiApoLp-IIIRNAi crickets. These observations indicated that ApoLp-III does not maintain basal hemolymph lipid levels in crickets fed ad libitum, but is necessary for mobilizing lipid transfer into the hemolymph following AKH stimulation. Second, Lp (containing lipids) was injected into the hemolymph to induce a temporary increase in hemolymph lipid levels. Consequently, the initiation of feeding was delayed in a dose-dependent manner, indicating that increased hemolymph lipid levels reduced the motivation to feed. Taken together, these data validate the importance of basal hemolymph lipid levels in the control of energy homeostasis and for regulating feeding behavior in crickets.
Conflict of interest statement
Figures
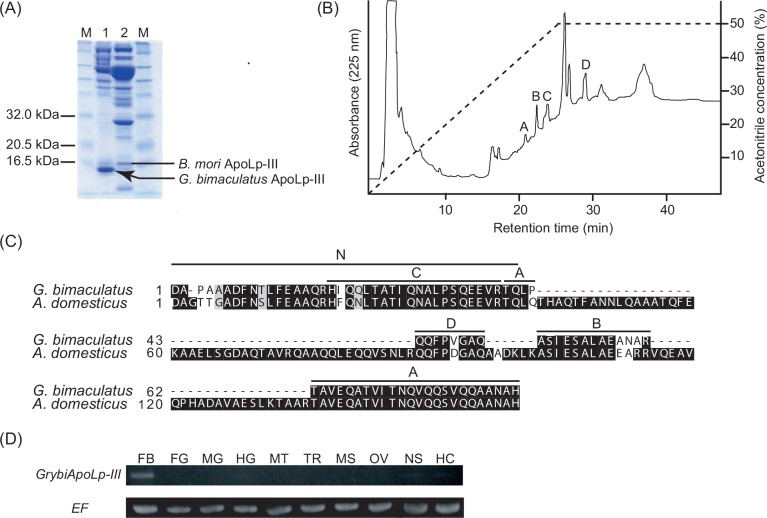
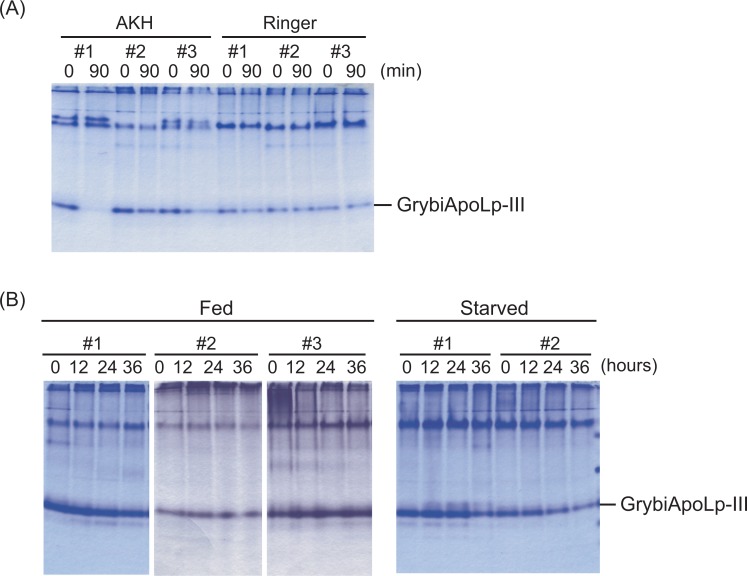
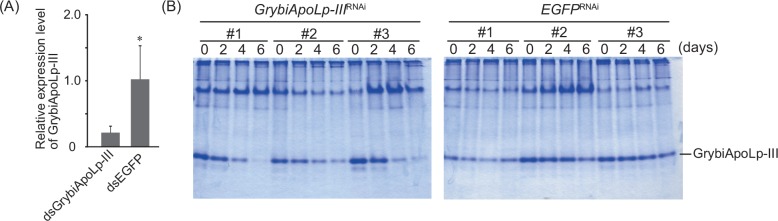
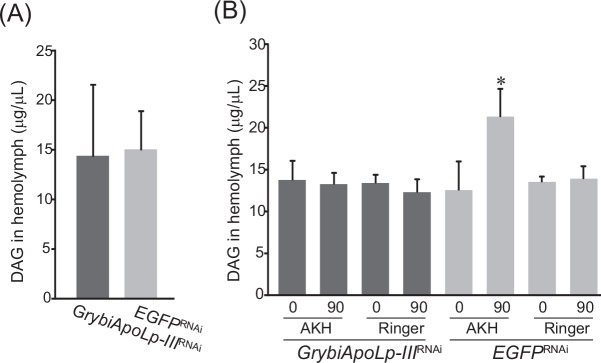
Similar articles
-
Adipokinetic hormone signaling determines dietary fatty acid preference through maintenance of hemolymph fatty acid composition in the cricket Gryllus bimaculatus.Sci Rep. 2018 Mar 16;8(1):4737. doi: 10.1038/s41598-018-22987-2. Sci Rep. 2018. PMID: 29549314 Free PMC article.
-
Effects of adipokinetic hormone and its related peptide on maintaining hemolymph carbohydrate and lipid levels in the two-spotted cricket, Gryllus bimaculatus.Biosci Biotechnol Biochem. 2018 Feb;82(2):274-284. doi: 10.1080/09168451.2017.1422106. Epub 2018 Jan 12. Biosci Biotechnol Biochem. 2018. PMID: 29325488
-
Age-dependent changes of fat body stores and the regulation of fat body lipid synthesis and mobilisation by adipokinetic hormone in the last larval instar of the cricket, Gryllus bimaculatus.J Insect Physiol. 2008 Oct-Nov;54(10-11):1404-12. doi: 10.1016/j.jinsphys.2008.08.001. Epub 2008 Aug 7. J Insect Physiol. 2008. PMID: 18761344
-
Lipid transport biochemistry and its role in energy production.Annu Rev Entomol. 2000;45:233-60. doi: 10.1146/annurev.ento.45.1.233. Annu Rev Entomol. 2000. PMID: 10761577 Review.
-
Insect adipokinetic hormones: release and integration of flight energy metabolism.Comp Biochem Physiol B Biochem Mol Biol. 2003 Oct;136(2):217-26. doi: 10.1016/s1096-4959(03)00151-9. Comp Biochem Physiol B Biochem Mol Biol. 2003. PMID: 14529748 Review.
Cited by
-
Drinking Warm Water Promotes Performance by Regulating Ruminal Microbial Composition and Serum Metabolites in Yak Calves.Microorganisms. 2023 Aug 16;11(8):2092. doi: 10.3390/microorganisms11082092. Microorganisms. 2023. PMID: 37630652 Free PMC article.
-
Two neuropeptide signaling pathways regulate post-mating refractoriness and reproductive system in male crickets.Commun Biol. 2025 Jun 11;8(1):910. doi: 10.1038/s42003-025-08219-0. Commun Biol. 2025. PMID: 40500289 Free PMC article.
-
CCHamide-2 Signaling Regulates Food Intake and Metabolism in Gryllus bimaculatus.Insects. 2022 Mar 25;13(4):324. doi: 10.3390/insects13040324. Insects. 2022. PMID: 35447766 Free PMC article.
-
Adipokinetic hormone signaling determines dietary fatty acid preference through maintenance of hemolymph fatty acid composition in the cricket Gryllus bimaculatus.Sci Rep. 2018 Mar 16;8(1):4737. doi: 10.1038/s41598-018-22987-2. Sci Rep. 2018. PMID: 29549314 Free PMC article.
References
-
- Bernays EA, Chapman RF (1973) The regulation of feeding in Locusta migratoria: Internal inhibitory mechanisms. Entomol Exp Appl 16:329–342
-
- Simpson SJ (1981) An oscillation underlying feeding and a number of other behaviours in fifth-instar Locusta migratoria nymphs. Physiol Entomol 6:315–324
-
- Simpson SJ, Ludlow AR (1986) Why locusts start to feed: A comparison of causal factors. Anim Behav 34:480–496
-
- Simpson SJ (1995) The control of meals in chewing insects In:Chapman RF, de Boer G., (Eds.), Regulatory Mechanisms of Insect Feeding, Chapman and Hall, New York, pp. 137–156
-
- Simpson SJ (1983) The role of volumetric feedback from the hindgut in the regulation of meal size in fifth-instar Locusta migratoria nymphs. Physiol Entomol 8:451–467
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
