

Enteroviral proteases: structure, host interactions and pathogenicity
- PMID: 27145174
- PMCID: PMC7169145
- DOI: 10.1002/rmv.1883
Enteroviral proteases: structure, host interactions and pathogenicity
Abstract
Enteroviruses are common human pathogens, and infections are particularly frequent in children. Severe infections can lead to a variety of diseases, including poliomyelitis, aseptic meningitis, myocarditis and neonatal sepsis. Enterovirus infections have also been implicated in asthmatic exacerbations and type 1 diabetes. The large disease spectrum of the closely related enteroviruses may be partially, but not fully, explained by differences in tissue tropism. The molecular mechanisms by which enteroviruses cause disease are poorly understood, but there is increasing evidence that the two enteroviral proteases, 2A(pro) and 3C(pro) , are important mediators of pathology. These proteases perform the post-translational proteolytic processing of the viral polyprotein, but they also cleave several host-cell proteins in order to promote the production of new virus particles, as well as to evade the cellular antiviral immune responses. Enterovirus-associated processing of cellular proteins may also contribute to pathology, as elegantly demonstrated by the 2A(pro) -mediated cleavage of dystrophin in cardiomyocytes contributing to Coxsackievirus-induced cardiomyopathy. It is likely that improved tools to identify targets for these proteases will reveal additional host protein substrates that can be linked to specific enterovirus-associated diseases. Here, we discuss the function of the enteroviral proteases in the virus replication cycle and review the current knowledge regarding how these proteases modulate the infected cell in order to favour virus replication, including ways to avoid detection by the immune system. We also highlight new possibilities for the identification of protease-specific cellular targets and thereby a way to discover novel mechanisms contributing to disease. Copyright © 2016 John Wiley & Sons, Ltd.
Copyright © 2016 John Wiley & Sons, Ltd.
Conflict of interest statement
The authors have no competing interests.
Figures


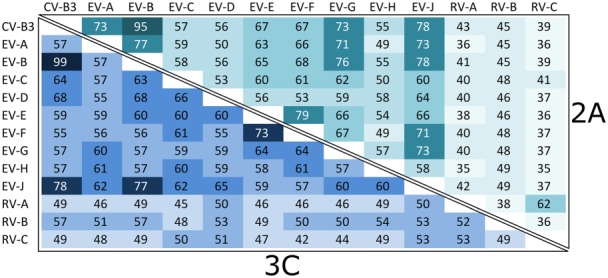
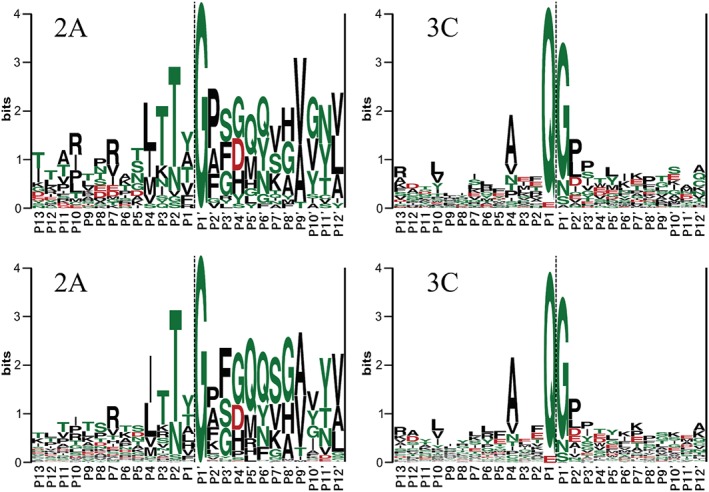
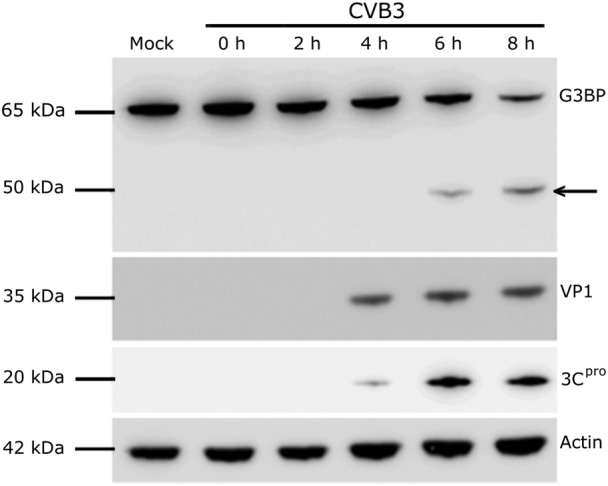
References
-
- Tapparel C, Siegrist F, Petty TJ, et al. Picornavirus and enterovirus diversity with associated human diseases. Infection, Genetics and Evolution 2013; 14: 282–293. - PubMed
-
- Hober D, Alidjinou EK. Enteroviral pathogenesis of type 1 diabetes: queries and answers. Current Opinion in Infectious Diseases 2013; 26: 263–269. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
