

Correlations and Neuronal Population Information
- PMID: 27145916
- PMCID: PMC5137197
- DOI: 10.1146/annurev-neuro-070815-013851
Correlations and Neuronal Population Information
Abstract
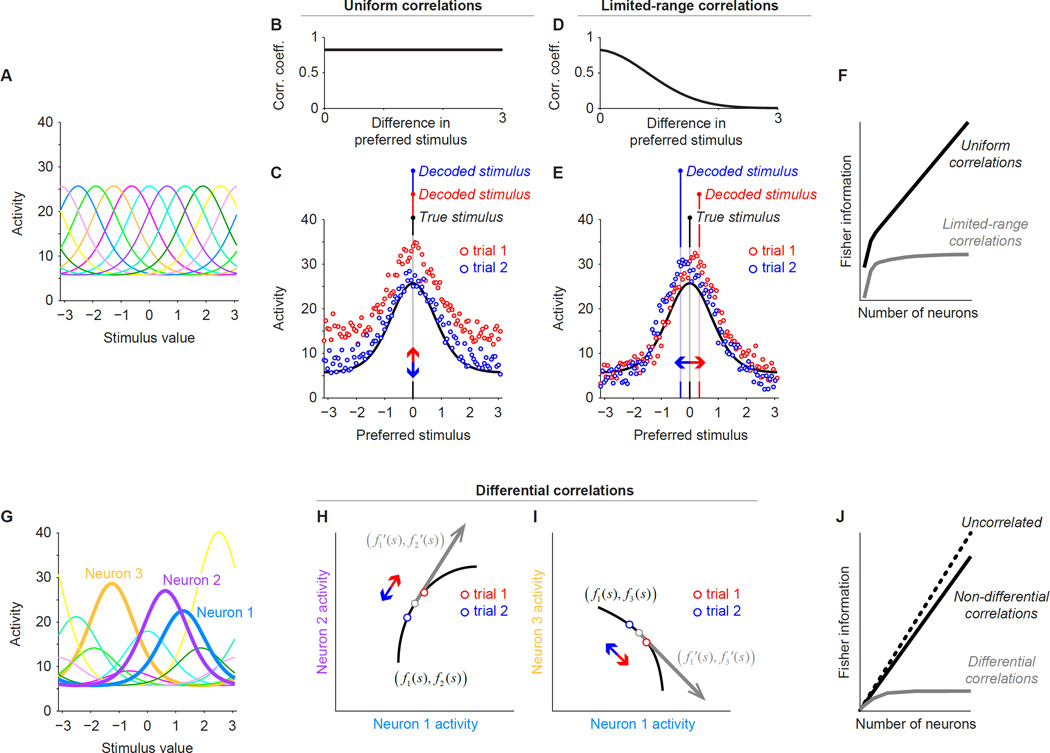
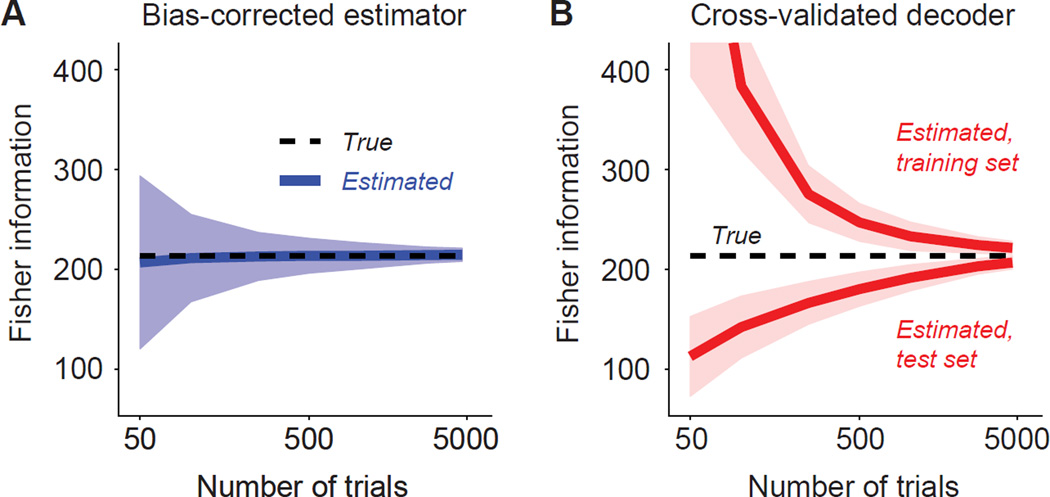
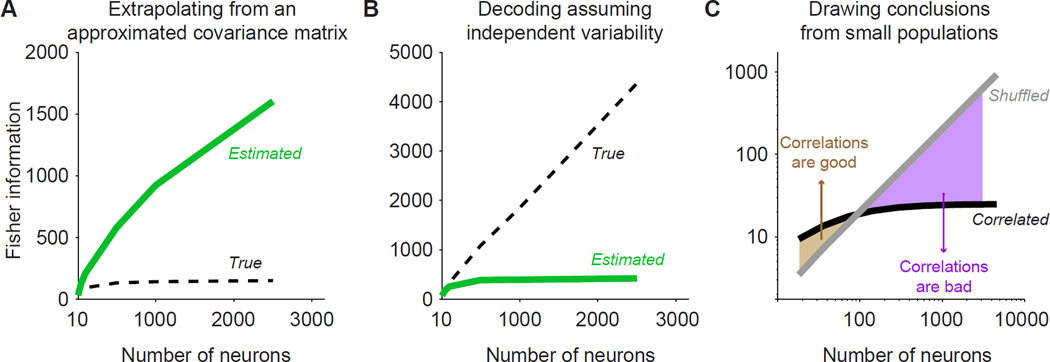
Brain function involves the activity of neuronal populations. Much recent effort has been devoted to measuring the activity of neuronal populations in different parts of the brain under various experimental conditions. Population activity patterns contain rich structure, yet many studies have focused on measuring pairwise relationships between members of a larger population-termed noise correlations. Here we review recent progress in understanding how these correlations affect population information, how information should be quantified, and what mechanisms may give rise to correlations. As population coding theory has improved, it has made clear that some forms of correlation are more important for information than others. We argue that this is a critical lesson for those interested in neuronal population responses more generally: Descriptions of population responses should be motivated by and linked to well-specified function. Within this context, we offer suggestions of where current theoretical frameworks fall short.
Keywords: Fisher information; decoding; neural coding; neural variability; perception; theoretical neuroscience.
Figures
References
-
- Abbott LF, Dayan P. The effect of correlated variability on the accuracy of a population code. Neural Comput. 1999;11:91–101. - PubMed
-
- Arieli A, Sterkin A, Grinvald A, Aertsen A. Dynamics of ongoing activity: explanation of the large variability in evoked cortical responses. Science. 1996;273:1868–1871. - PubMed
-
- Atick J, Redlich A. Towards a theory of early visual processing. Neural Comput. 1990;2:308–320.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
