

How Bacteria Subvert Animal Cell Structure and Function
- PMID: 27146312
- PMCID: PMC5178824
- DOI: 10.1146/annurev-cellbio-100814-125227
How Bacteria Subvert Animal Cell Structure and Function
Abstract
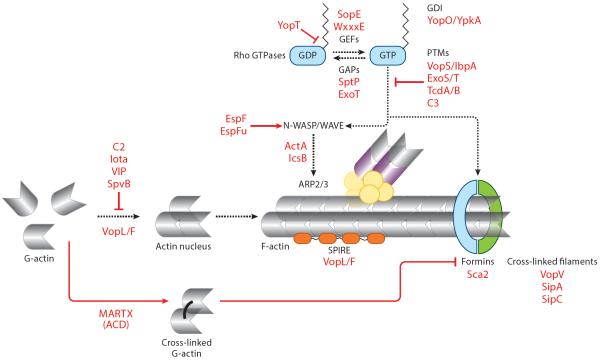
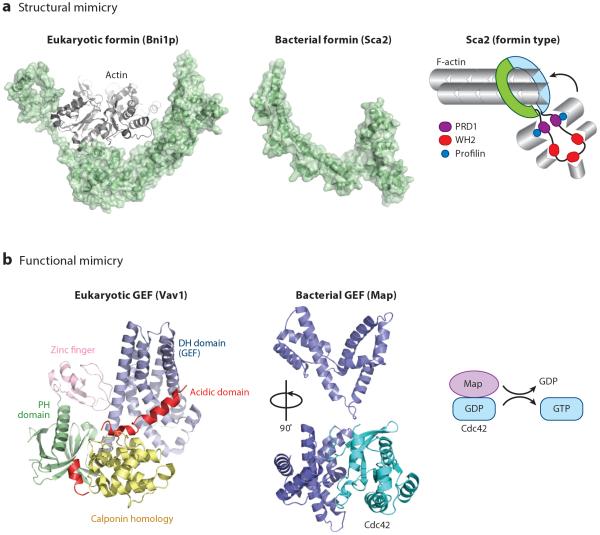
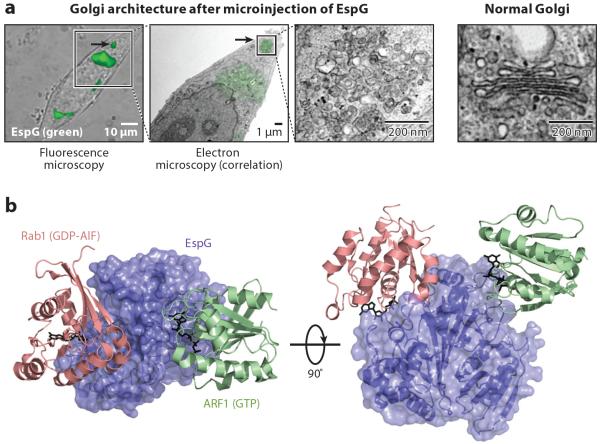
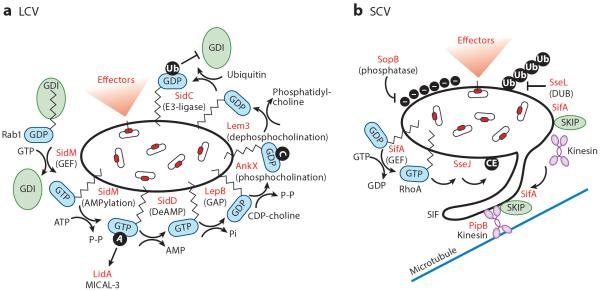
Bacterial pathogens encode a wide variety of effectors and toxins that hijack host cell structure and function. Of particular importance are virulence factors that target actin cytoskeleton dynamics critical for cell shape, stability, motility, phagocytosis, and division. In addition, many bacteria target organelles of the general secretory pathway (e.g., the endoplasmic reticulum and the Golgi complex) and recycling pathways (e.g., the endolysosomal system) to establish and maintain an intracellular replicative niche. Recent research on the biochemistry and structural biology of bacterial effector proteins and toxins has begun to shed light on the molecular underpinnings of these host-pathogen interactions. This exciting work is revealing how pathogens gain control of the complex and dynamic host cellular environments, which impacts our understanding of microbial infectious disease, immunology, and human cell biology.
Keywords: Golgi complex; actin cytoskeleton; bacterial effectors; bacterial toxins; endosomal/lysosomal trafficking.
Figures
References
-
- Aili M, Hallberg B, Wolf-Watz H, Rosqvist R. GAP activity of Yersinia YopE. Methods Enzymol. 2002;358:359–70. - PubMed
-
- Aktories K. Bacterial protein toxins that modify host regulatory GTPases. Nat. Rev. Microbiol. 2011;9:487–98. - PubMed
-
- Aktories K, Barmann M, Ohishi I, Tsuyama S, Jakobs KH, Habermann E. Botulinum C2 toxin ADP-ribosylates actin. Nature. 1986;322:390–92. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
