

An ancient yet flexible cis-regulatory architecture allows localized Hedgehog tuning by patched/Ptch1
- PMID: 27146892
- PMCID: PMC4887206
- DOI: 10.7554/eLife.13550
An ancient yet flexible cis-regulatory architecture allows localized Hedgehog tuning by patched/Ptch1
Abstract
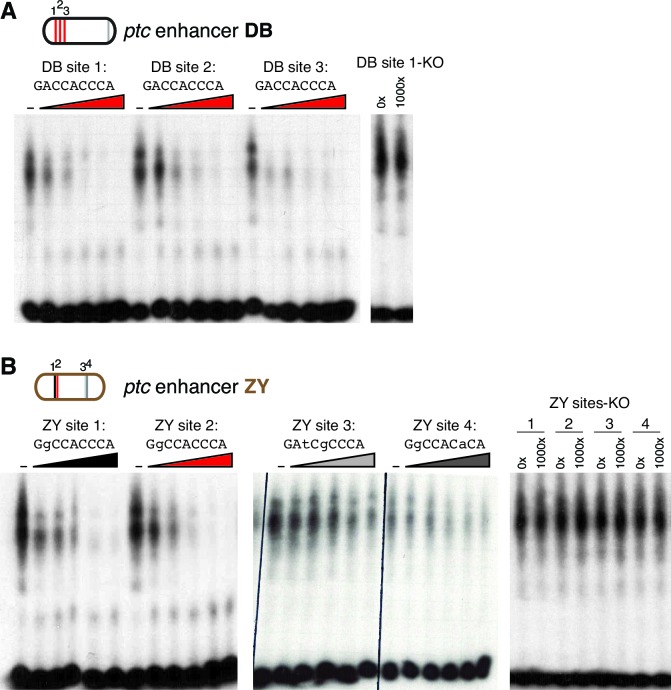
The Hedgehog signaling pathway is part of the ancient developmental-evolutionary animal toolkit. Frequently co-opted to pattern new structures, the pathway is conserved among eumetazoans yet flexible and pleiotropic in its effects. The Hedgehog receptor, Patched, is transcriptionally activated by Hedgehog, providing essential negative feedback in all tissues. Our locus-wide dissections of the cis-regulatory landscapes of fly patched and mouse Ptch1 reveal abundant, diverse enhancers with stage- and tissue-specific expression patterns. The seemingly simple, constitutive Hedgehog response of patched/Ptch1 is driven by a complex regulatory architecture, with batteries of context-specific enhancers engaged in promoter-specific interactions to tune signaling individually in each tissue, without disturbing patterning elsewhere. This structure-one of the oldest cis-regulatory features discovered in animal genomes-explains how patched/Ptch1 can drive dramatic adaptations in animal morphology while maintaining its essential core function. It may also suggest a general model for the evolutionary flexibility of conserved regulators and pathways.
Keywords: D. melanogaster; Hedgehog signaling; cis-regulatory elements; development; developmental biology; evolution of development (evo-devo); evolutionary biology; gene regulation; genomics; mouse; stem cells; transcriptional regulation.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures















Similar articles
-
Activities of the Cytoplasmic Domains of Patched-1 Modulate but Are Not Essential for the Regulation of Canonical Hedgehog Signaling.J Biol Chem. 2016 Aug 19;291(34):17557-68. doi: 10.1074/jbc.M116.731745. Epub 2016 Jun 20. J Biol Chem. 2016. PMID: 27325696 Free PMC article.
-
Structural Basis for Cholesterol Transport-like Activity of the Hedgehog Receptor Patched.Cell. 2018 Nov 15;175(5):1352-1364.e14. doi: 10.1016/j.cell.2018.10.026. Epub 2018 Nov 8. Cell. 2018. PMID: 30415841 Free PMC article.
-
Ptch2/Gas1 and Ptch1/Boc differentially regulate Hedgehog signalling in murine primordial germ cell migration.Nat Commun. 2020 Apr 24;11(1):1994. doi: 10.1038/s41467-020-15897-3. Nat Commun. 2020. PMID: 32332736 Free PMC article.
-
Hedgehog signaling pathway and gastrointestinal stem cell signaling network (review).Int J Mol Med. 2006 Dec;18(6):1019-23. Int J Mol Med. 2006. PMID: 17089004 Review.
-
Drosophila Segment Polarity Mutants and the Rediscovery of the Hedgehog Pathway Genes.Curr Top Dev Biol. 2016;116:477-88. doi: 10.1016/bs.ctdb.2016.01.007. Epub 2016 Feb 13. Curr Top Dev Biol. 2016. PMID: 26970635 Review.
Cited by
-
The formation of the thumb requires direct modulation of Gli3 transcription by Hoxa13.Proc Natl Acad Sci U S A. 2020 Jan 14;117(2):1090-1096. doi: 10.1073/pnas.1919470117. Epub 2020 Jan 2. Proc Natl Acad Sci U S A. 2020. PMID: 31896583 Free PMC article.
-
MicroRNA-129-5p suppresses nasopharyngeal carcinoma lymphangiogenesis and lymph node metastasis by targeting ZIC2.Cell Oncol (Dordr). 2020 Apr;43(2):249-261. doi: 10.1007/s13402-019-00485-5. Epub 2019 Dec 28. Cell Oncol (Dordr). 2020. PMID: 31884576
-
Atlas of nascent RNA transcripts reveals enhancer to gene linkages.bioRxiv [Preprint]. 2023 Dec 8:2023.12.07.570626. doi: 10.1101/2023.12.07.570626. bioRxiv. 2023. Update in: BMC Genomics. 2025 Apr 25;26(1):406. doi: 10.1186/s12864-025-11568-z. PMID: 38105978 Free PMC article. Updated. Preprint.
-
Regulation of GLI1 by cis DNA elements and epigenetic marks.DNA Repair (Amst). 2019 Jul;79:10-21. doi: 10.1016/j.dnarep.2019.04.011. Epub 2019 May 2. DNA Repair (Amst). 2019. PMID: 31085420 Free PMC article.
-
Tuned polymerization of the transcription factor Yan limits off-DNA sequestration to confer context-specific repression.Elife. 2018 Nov 9;7:e37545. doi: 10.7554/eLife.37545. Elife. 2018. PMID: 30412049 Free PMC article.
References
-
- Alexandre C, Jacinto A, Ingham PW. Transcriptional activation of hedgehog target genes in Drosophila is mediated directly by the cubitus interruptus protein, a member of the GLI family of zinc finger DNA-binding proteins. Genes & Development. 1996;10:2003–2013. doi: 10.1101/gad.10.16.2003. - DOI - PubMed
-
- Barolo S, Carver LA, Posakony JW. GFP and beta-galactosidase transformation vectors for promoter/enhancer analysis in Drosophila. BioTechniques. 2000;29 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
