

Characterization of a Mutant Deficient for Ammonium and Nitric Oxide Signalling in the Model System Chlamydomonas reinhardtii
- PMID: 27149516
- PMCID: PMC4858171
- DOI: 10.1371/journal.pone.0155128
Characterization of a Mutant Deficient for Ammonium and Nitric Oxide Signalling in the Model System Chlamydomonas reinhardtii
Abstract
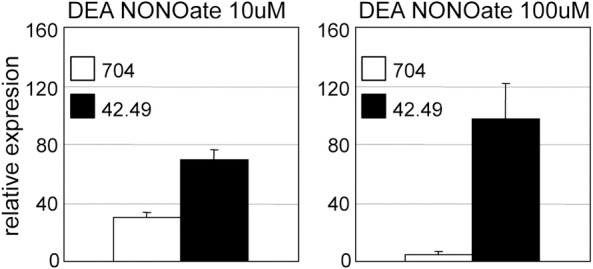
The ubiquitous signalling molecule Nitric Oxide (NO) is characterized not only by the variety of organisms in which it has been described, but also by the wealth of biological processes that it regulates. In contrast to the expanding repertoire of functions assigned to NO, however, the mechanisms of NO action usually remain unresolved, and genes that work within NO signalling cascades are seldom identified. A recent addition to the list of known NO functions is the regulation of the nitrogen assimilation pathway in the unicellular alga Chlamydomonas reinhardtii, a well-established model organism for genetic and molecular studies that offers new possibilities in the search for mediators of NO signalling. By further exploiting a collection of Chlamydomonas insertional mutant strains originally isolated for their insensitivity to the ammonium (NH4+) nitrogen source, we found a mutant which, in addition to its ammonium insensitive (AI) phenotype, was not capable of correctly sensing the NO signal. Similarly to what had previously been described in the AI strain cyg56, the expression of nitrogen assimilation genes in the mutant did not properly respond to treatments with various NO donors. Complementation experiments showed that NON1 (NO Nitrate 1), a gene that encodes a protein containing no known functional domain, was the gene underlying the mutant phenotype. Beyond the identification of NON1, our findings broadly demonstrate the potential for Chlamydomonas reinhardtii to be used as a model system in the search for novel components of gene networks that mediate physiological responses to NO.
Conflict of interest statement
Figures
Similar articles
-
Arginine is a component of the ammonium-CYG56 signalling cascade that represses genes of the nitrogen assimilation pathway in Chlamydomonas reinhardtii.PLoS One. 2018 Apr 23;13(4):e0196167. doi: 10.1371/journal.pone.0196167. eCollection 2018. PLoS One. 2018. PMID: 29684072 Free PMC article.
-
Transcriptional regulation of CDP1 and CYG56 is required for proper NH4+ sensing in Chlamydomonas.J Exp Bot. 2011 Feb;62(4):1425-37. doi: 10.1093/jxb/erq384. Epub 2010 Dec 1. J Exp Bot. 2011. PMID: 21127023
-
Isolation and characterization of two new negative regulatory mutants for nitrate assimilation in Chlamydomonas reinhardtii obtained by insertional mutagenesis.Mol Gen Genet. 1996 Jun 24;251(4):461-71. doi: 10.1007/BF02172375. Mol Gen Genet. 1996. PMID: 8709950
-
How Chlamydomonas handles nitrate and the nitric oxide cycle.J Exp Bot. 2017 May 1;68(10):2593-2602. doi: 10.1093/jxb/erw507. J Exp Bot. 2017. PMID: 28201747 Review.
-
Insertional mutagenesis as a tool to study genes/functions in Chlamydomonas.Adv Exp Med Biol. 2007;616:77-89. doi: 10.1007/978-0-387-75532-8_7. Adv Exp Med Biol. 2007. PMID: 18161492 Review.
Cited by
-
Ten years of algal biofuel and bioproducts: gains and pains.Planta. 2019 Jan;249(1):195-219. doi: 10.1007/s00425-018-3066-8. Epub 2019 Jan 2. Planta. 2019. PMID: 30603791 Review.
-
Arginine is a component of the ammonium-CYG56 signalling cascade that represses genes of the nitrogen assimilation pathway in Chlamydomonas reinhardtii.PLoS One. 2018 Apr 23;13(4):e0196167. doi: 10.1371/journal.pone.0196167. eCollection 2018. PLoS One. 2018. PMID: 29684072 Free PMC article.
-
Truncated hemoglobin 1 is a new player in Chlamydomonas reinhardtii acclimation to sulfur deprivation.PLoS One. 2017 Oct 19;12(10):e0186851. doi: 10.1371/journal.pone.0186851. eCollection 2017. PLoS One. 2017. PMID: 29049377 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
