

Single-molecule decoding of combinatorially modified nucleosomes
- PMID: 27151869
- PMCID: PMC4904710
- DOI: 10.1126/science.aad7701
Single-molecule decoding of combinatorially modified nucleosomes
Abstract
Different combinations of histone modifications have been proposed to signal distinct gene regulatory functions, but this area is poorly addressed by existing technologies. We applied high-throughput single-molecule imaging to decode combinatorial modifications on millions of individual nucleosomes from pluripotent stem cells and lineage-committed cells. We identified definitively bivalent nucleosomes with concomitant repressive and activating marks, as well as other combinatorial modification states whose prevalence varies with developmental potency. We showed that genetic and chemical perturbations of chromatin enzymes preferentially affect nucleosomes harboring specific modification states. Last, we combined this proteomic platform with single-molecule DNA sequencing technology to simultaneously determine the modification states and genomic positions of individual nucleosomes. This single-molecule technology has the potential to address fundamental questions in chromatin biology and epigenetic regulation.
Copyright © 2016, American Association for the Advancement of Science.
Figures
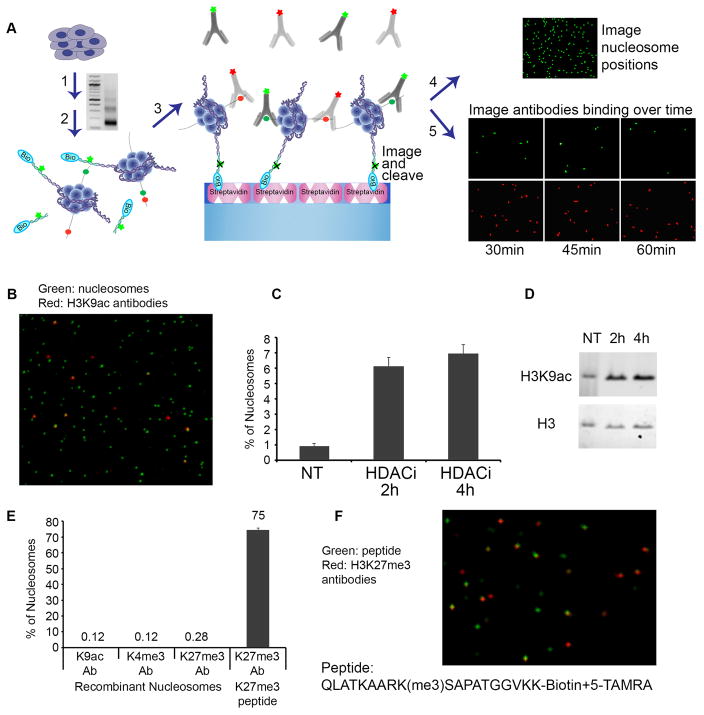
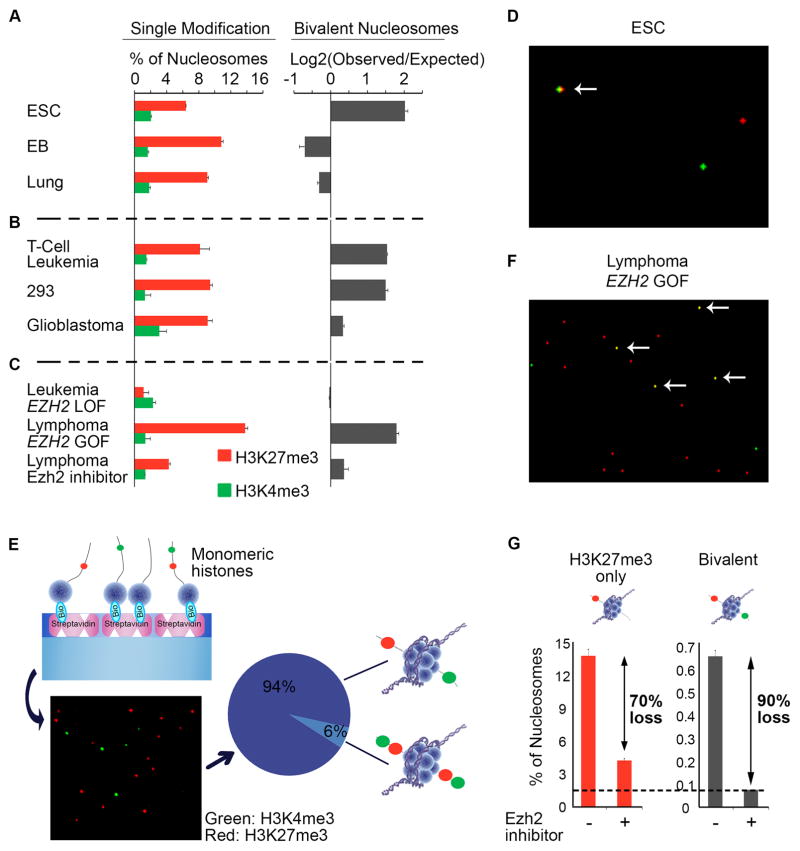
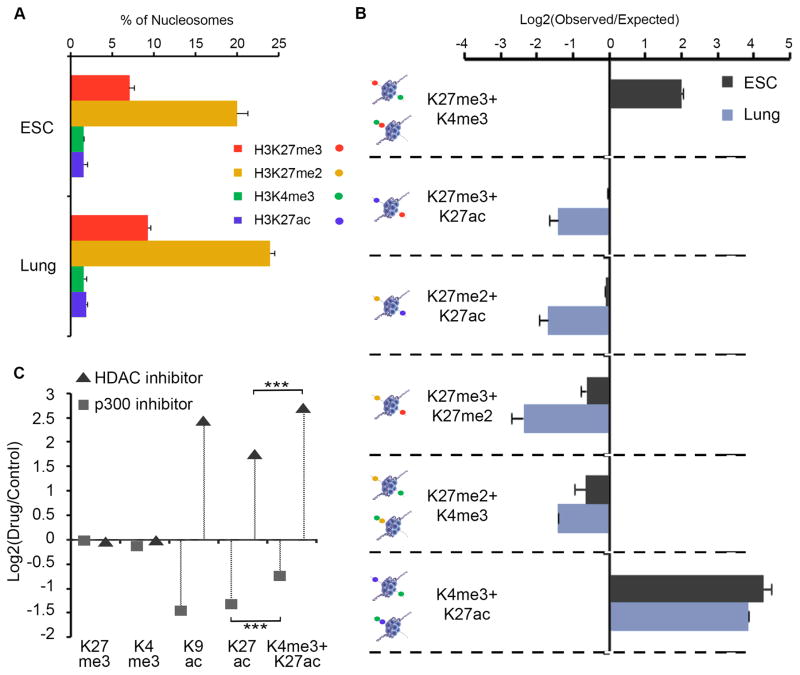
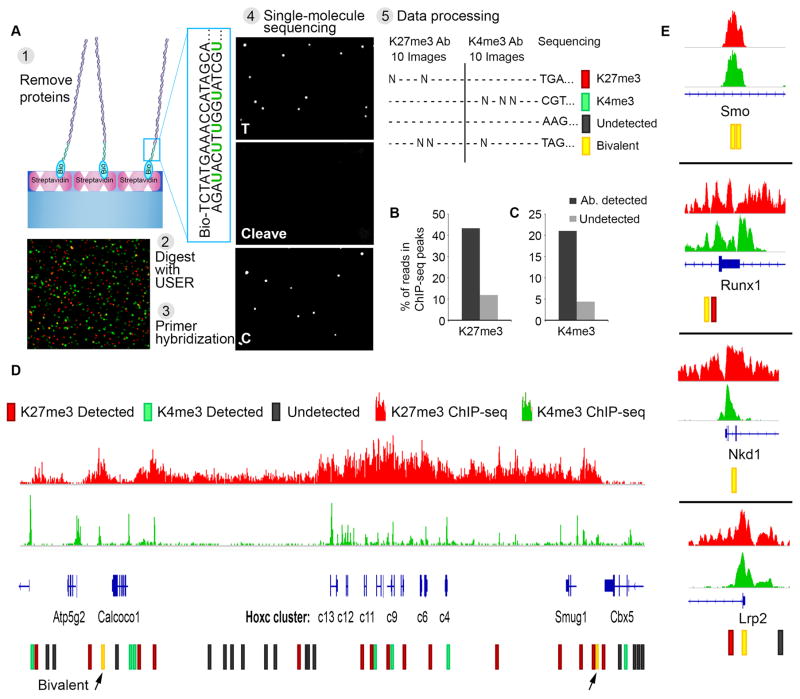
Comment in
-
Chromatin: Cracking the nucleosome code.Nat Rev Genet. 2016 Jul;17(7):376-7. doi: 10.1038/nrg.2016.70. Epub 2016 May 23. Nat Rev Genet. 2016. PMID: 27211067 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
