

Shape remodeling and blebbing of active cytoskeletal vesicles
- PMID: 27152328
- PMCID: PMC4846454
- DOI: 10.1126/sciadv.1500465
Shape remodeling and blebbing of active cytoskeletal vesicles
Abstract
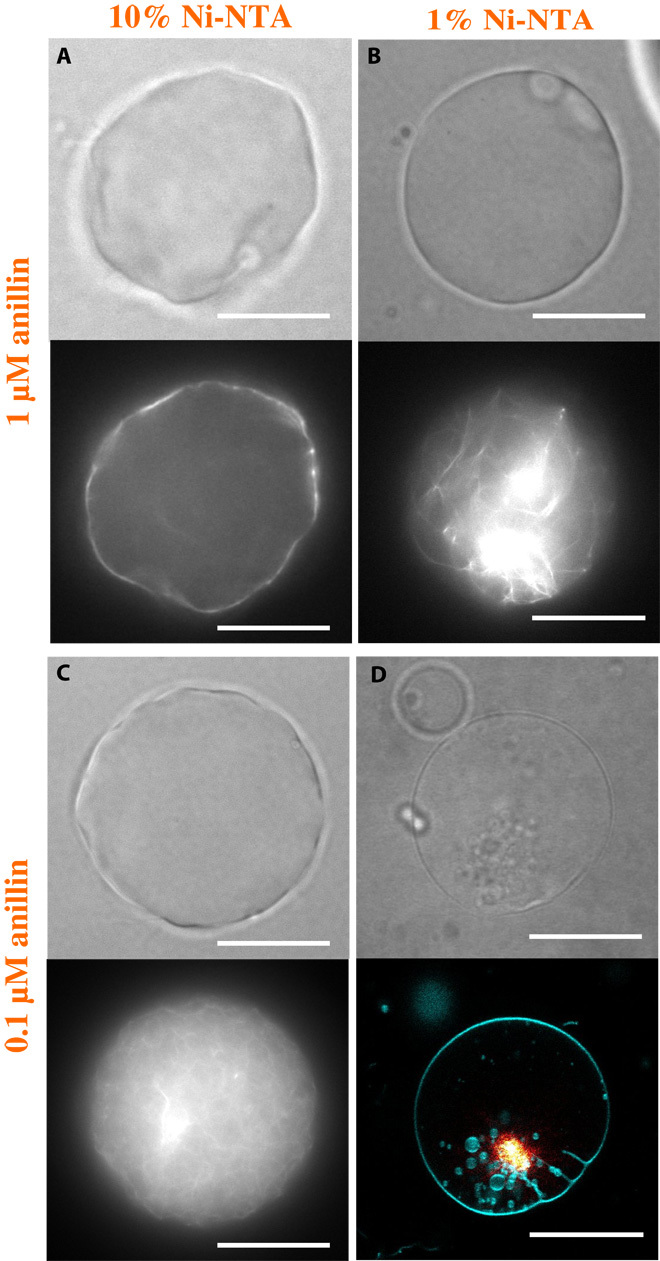
Morphological transformations of living cells, such as shape adaptation to external stimuli, blebbing, invagination, or tethering, result from an intricate interplay between the plasma membrane and its underlying cytoskeleton, where molecular motors generate forces. Cellular complexity defies a clear identification of the competing processes that lead to such a rich phenomenology. In a synthetic biology approach, designing a cell-like model assembled from a minimal set of purified building blocks would allow the control of all relevant parameters. We reconstruct actomyosin vesicles in which the coupling of the cytoskeleton to the membrane, the topology of the cytoskeletal network, and the contractile activity can all be precisely controlled and tuned. We demonstrate that tension generation of an encapsulated active actomyosin network suffices for global shape transformation of cell-sized lipid vesicles, which are reminiscent of morphological adaptations in living cells. The observed polymorphism of our cell-like model, such as blebbing, tether extrusion, or faceted shapes, can be qualitatively explained by the protein concentration dependencies and a force balance, taking into account the membrane tension, the density of anchoring points between the membrane and the actin network, and the forces exerted by molecular motors in the actin network. The identification of the physical mechanisms for shape transformations of active cytoskeletal vesicles sets a conceptual and quantitative benchmark for the further exploration of the adaptation mechanisms of cells.
Keywords: Active matter; actin networks; active cytoskeletal systems; biomimetic system; shape remodeling.
Figures




References
-
- Le Clainche C., Carlier M.-F., Regulation of actin assembly associated with protrusion and adhesion in cell migration. Physiol. Rev. 88, 489–513 (2008). - PubMed
-
- Lecuit T., Lenne P.-F., Munro E., Force generation, transmission, and integration during cell and tissue morphogenesis. Annu. Rev. Cell Dev. Biol. 27, 157–184 (2011). - PubMed
-
- Charras G., Paluch E., Blebs lead the way: How to migrate without lamellipodia. Nat. Rev. Mol. Cell Biol. 9, 730–736 (2008). - PubMed
-
- Döbereiner H.-G., Evans E., Kraus M., Seifert U., Wortis M., Mapping vesicle shapes into the phase diagram: A comparison of experiment and theory. Phys. Rev. E 55, 4458–4474 (1997).
-
- Berndl K., Käs J., Lipowsky R., Sackmann E., Seifert U., Shape transformations of giant vesicles: Extreme sensitivity to bilayer asymmetry. Europhys. Lett. 13, 659 (1990).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
