

Syntax compensates for poor binding sites to encode tissue specificity of developmental enhancers
- PMID: 27155014
- PMCID: PMC4988596
- DOI: 10.1073/pnas.1605085113
Syntax compensates for poor binding sites to encode tissue specificity of developmental enhancers
Abstract
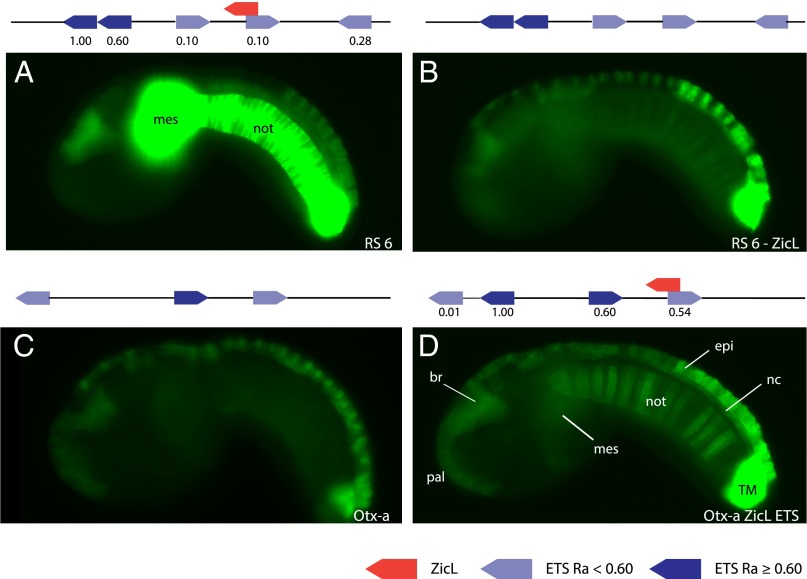
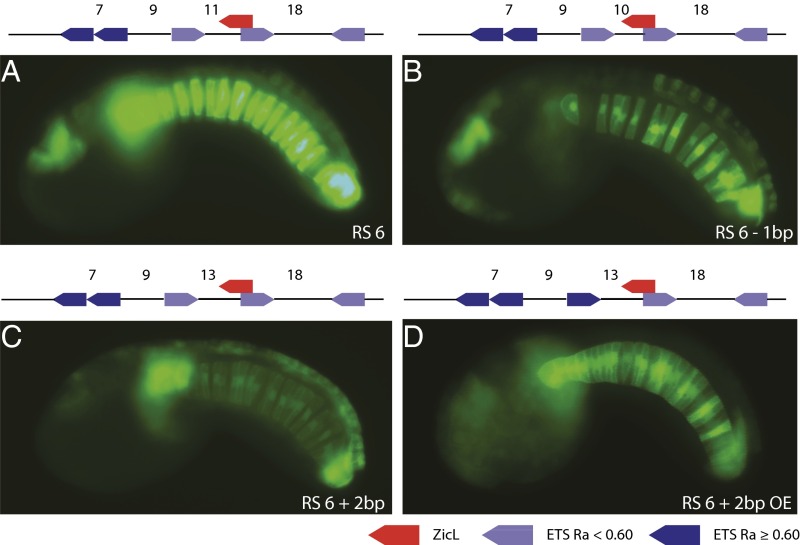
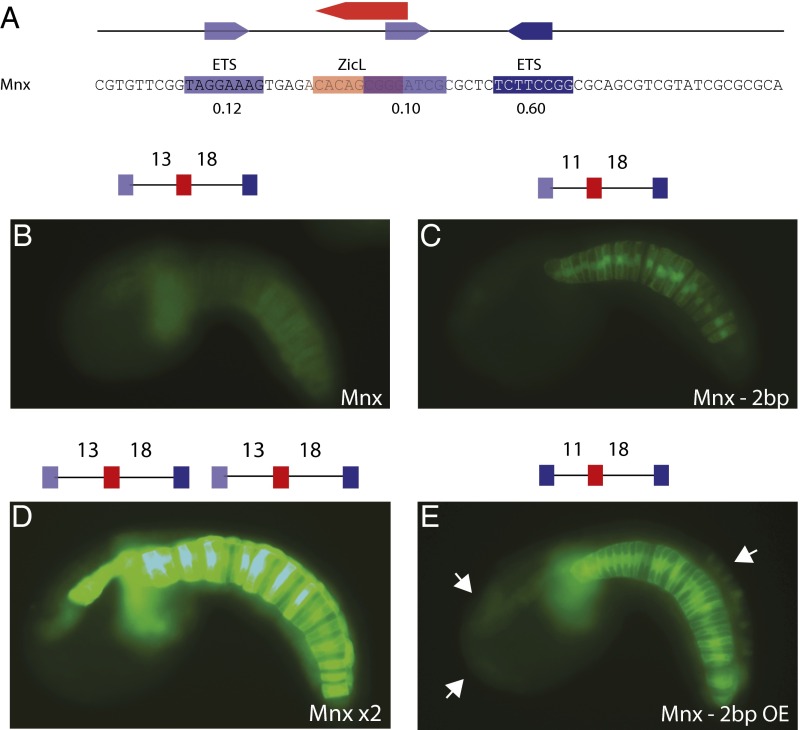
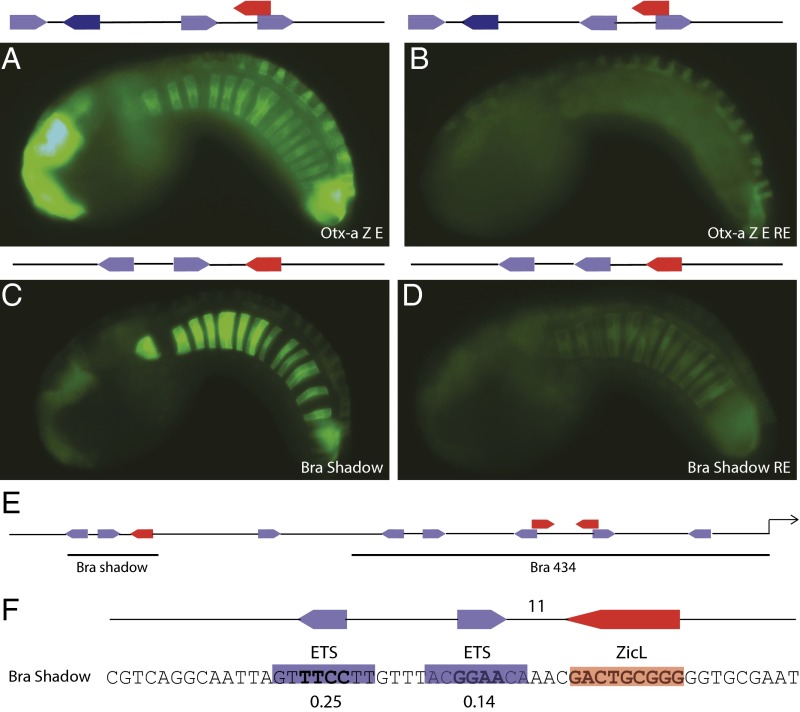
Transcriptional enhancers are short segments of DNA that switch genes on and off in response to a variety of intrinsic and extrinsic signals. Despite the discovery of the first enhancer more than 30 y ago, the relationship between primary DNA sequence and enhancer activity remains obscure. In particular, the importance of "syntax" (the order, orientation, and spacing of binding sites) is unclear. A high-throughput screen identified synthetic notochord enhancers that are activated by the combination of ZicL and ETS transcription factors in Ciona embryos. Manipulation of these enhancers elucidated a "regulatory code" of sequence and syntax features for notochord-specific expression. This code enabled in silico discovery of bona fide notochord enhancers, including those containing low-affinity binding sites that would be excluded by standard motif identification methods. One of the newly identified enhancers maps upstream of the known enhancer that regulates Brachyury (Ci-Bra), a key determinant of notochord specification. This newly identified Ci-Bra shadow enhancer contains binding sites with very low affinity, but optimal syntax, and therefore mediates surprisingly strong expression in the notochord. Weak binding sites are compensated by optimal syntax, whereas enhancers containing high-affinity binding affinities possess suboptimal syntax. We suggest this balance has obscured the importance of regulatory syntax, as noncanonical binding motifs are typically disregarded by enhancer detection methods. As a result, enhancers with low binding affinities but optimal syntax may be a vastly underappreciated feature of the regulatory genome.
Keywords: enhancer; enhancer grammar; gene regulation; regulatory principles; transcription.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Regulatory elements: Putting enhancers into context.Nat Rev Genet. 2016 Jul;17(7):377. doi: 10.1038/nrg.2016.74. Epub 2016 May 31. Nat Rev Genet. 2016. PMID: 27240812 No abstract available.
-
How to tune an enhancer.Proc Natl Acad Sci U S A. 2016 Jun 7;113(23):6330-1. doi: 10.1073/pnas.1606109113. Epub 2016 May 31. Proc Natl Acad Sci U S A. 2016. PMID: 27247414 Free PMC article. No abstract available.
Comment on
-
How to tune an enhancer.Proc Natl Acad Sci U S A. 2016 Jun 7;113(23):6330-1. doi: 10.1073/pnas.1606109113. Epub 2016 May 31. Proc Natl Acad Sci U S A. 2016. PMID: 27247414 Free PMC article. No abstract available.
References
-
- Small S, Kraut R, Hoey T, Warrior R, Levine M. Transcriptional regulation of a pair-rule stripe in Drosophila. Genes Dev. 1991;5(5):827–839. - PubMed
-
- Jiang J, Levine M. Binding affinities and cooperative interactions with bHLH activators delimit threshold responses to the dorsal gradient morphogen. Cell. 1993;72(5):741–752. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
