

Developmental Acquisition of Regulomes Underlies Innate Lymphoid Cell Functionality
- PMID: 27156451
- PMCID: PMC4874839
- DOI: 10.1016/j.cell.2016.04.029
Developmental Acquisition of Regulomes Underlies Innate Lymphoid Cell Functionality
Abstract
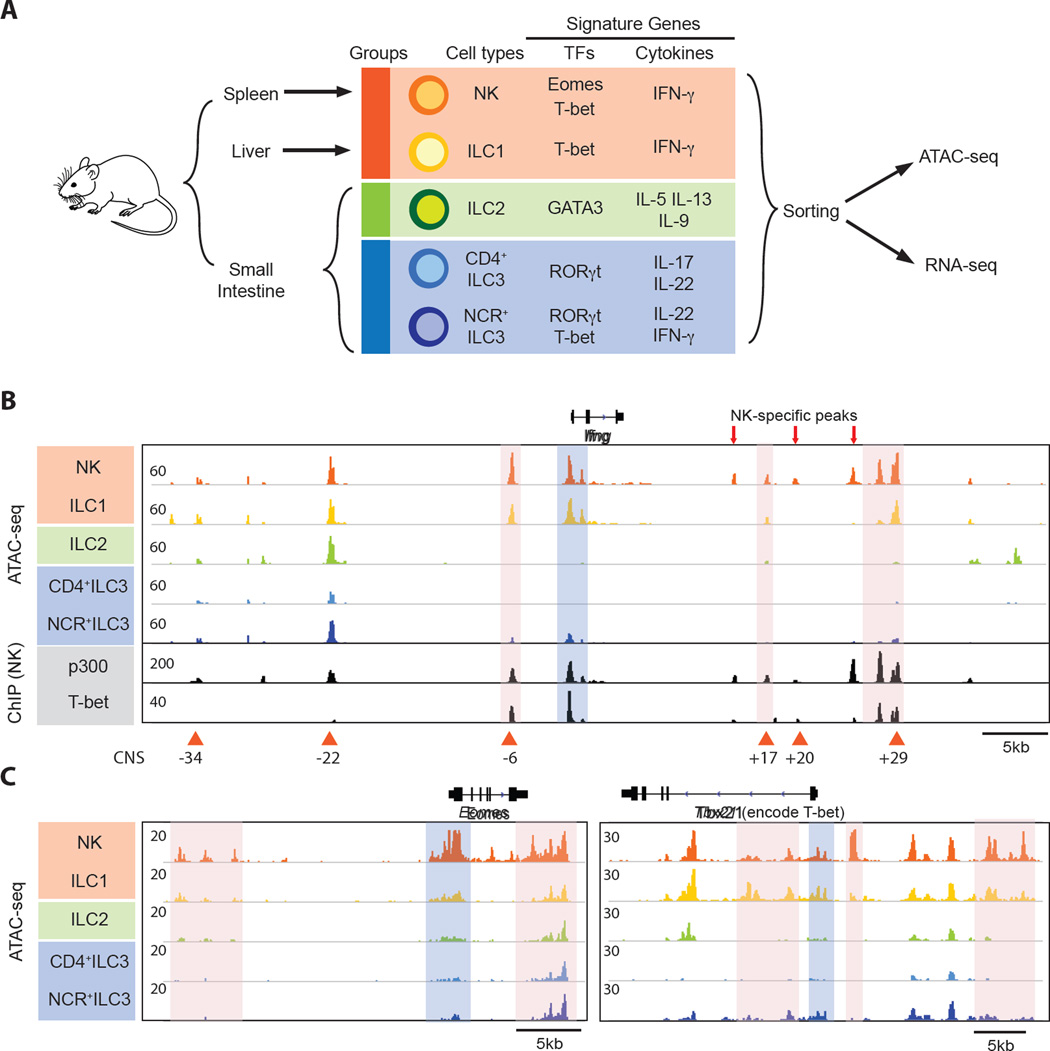
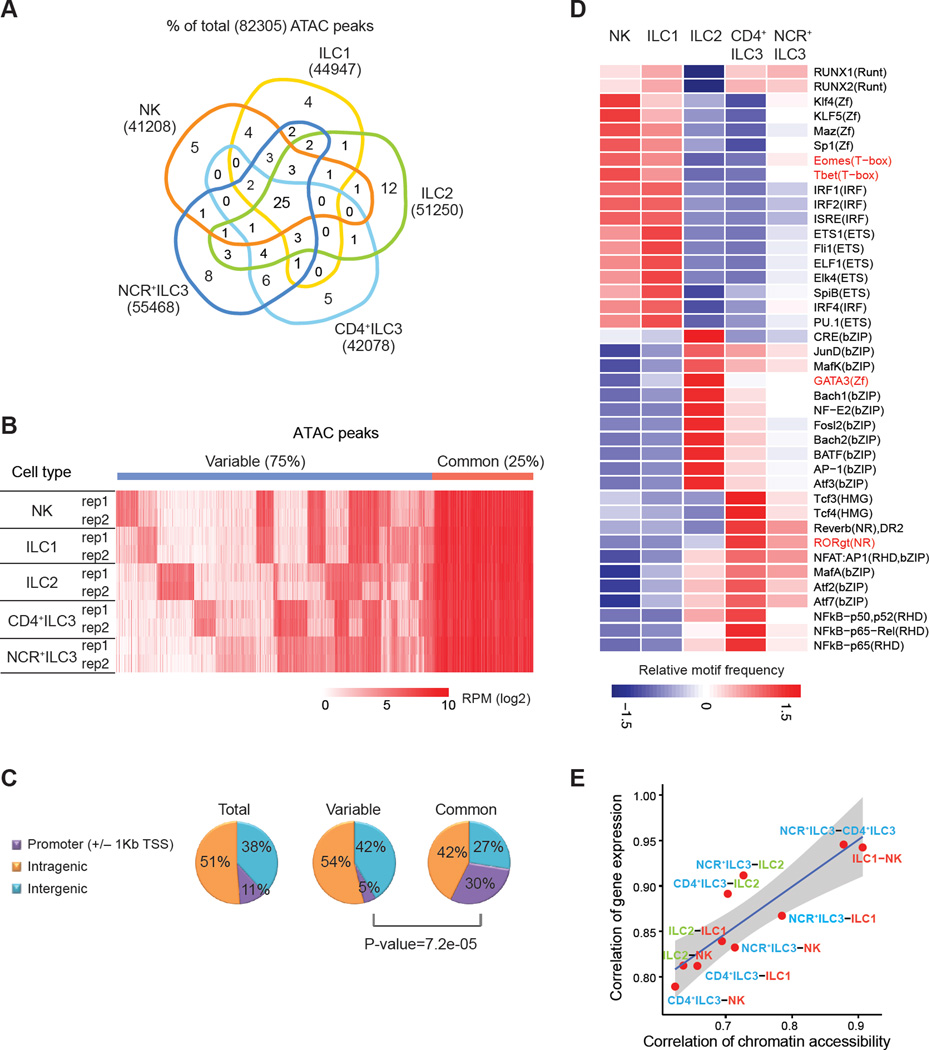
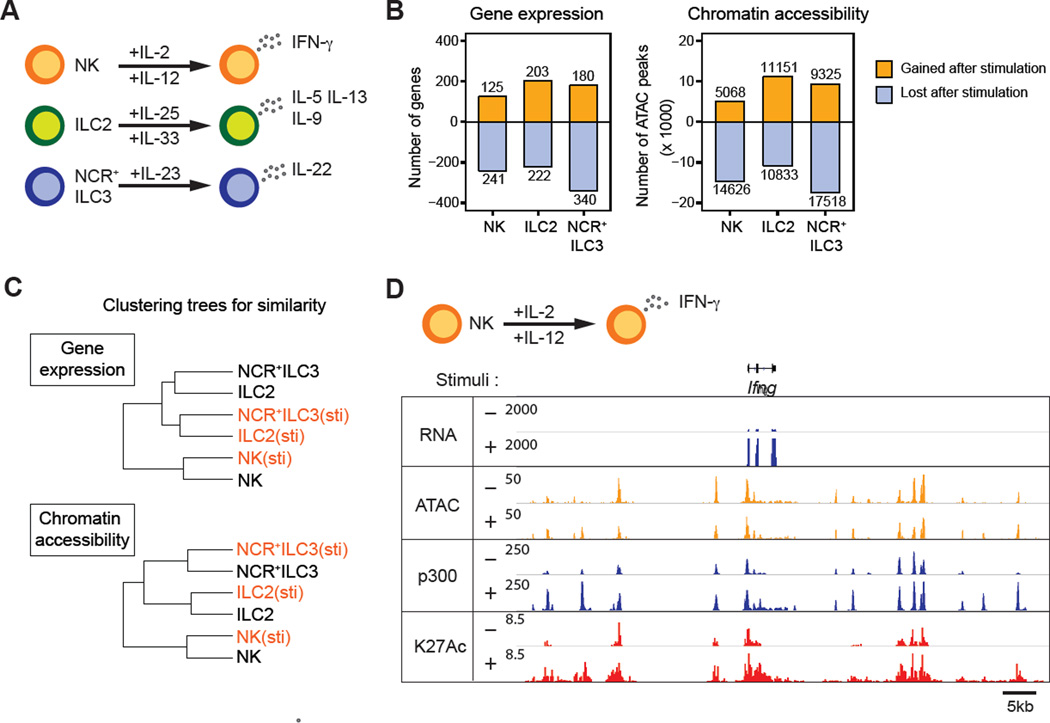
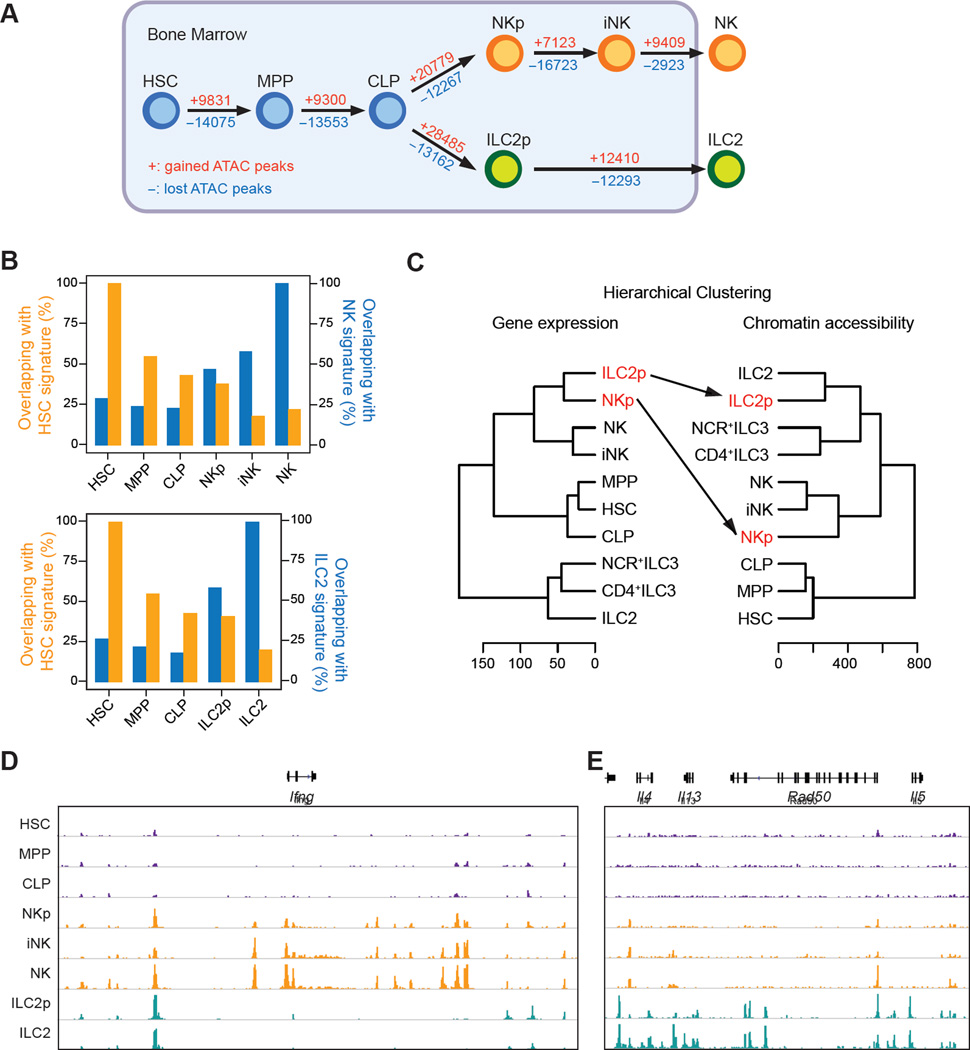
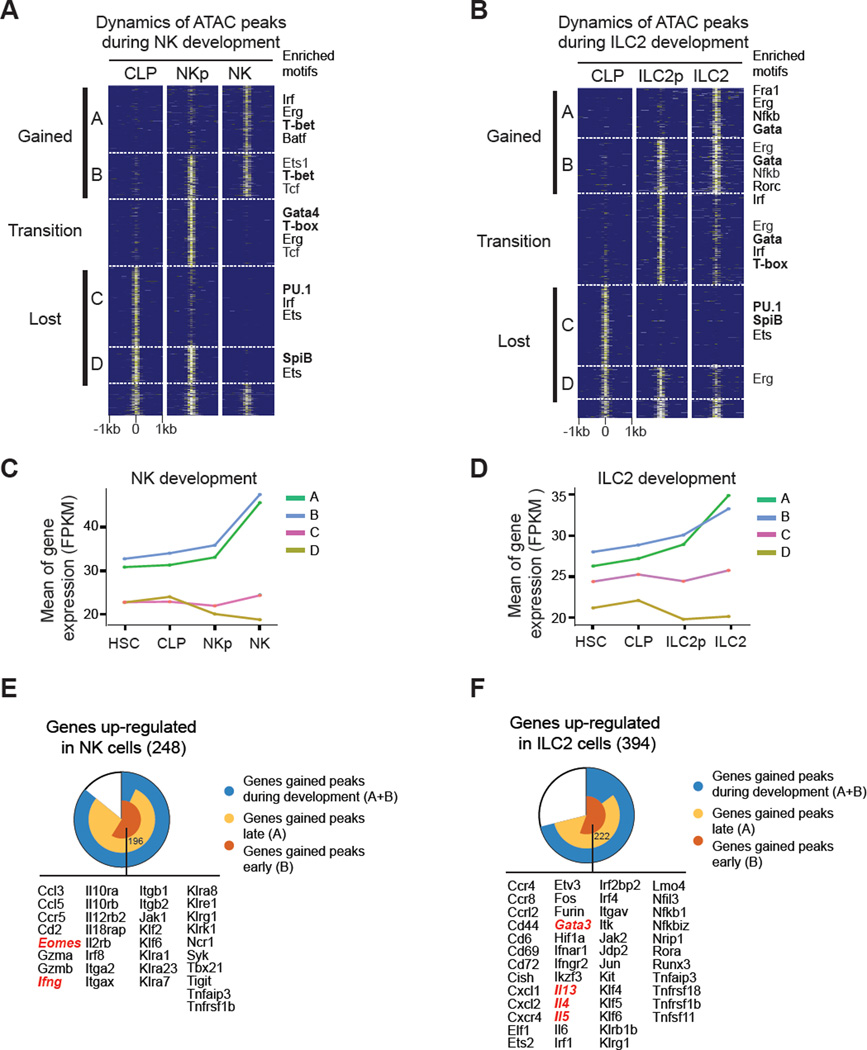
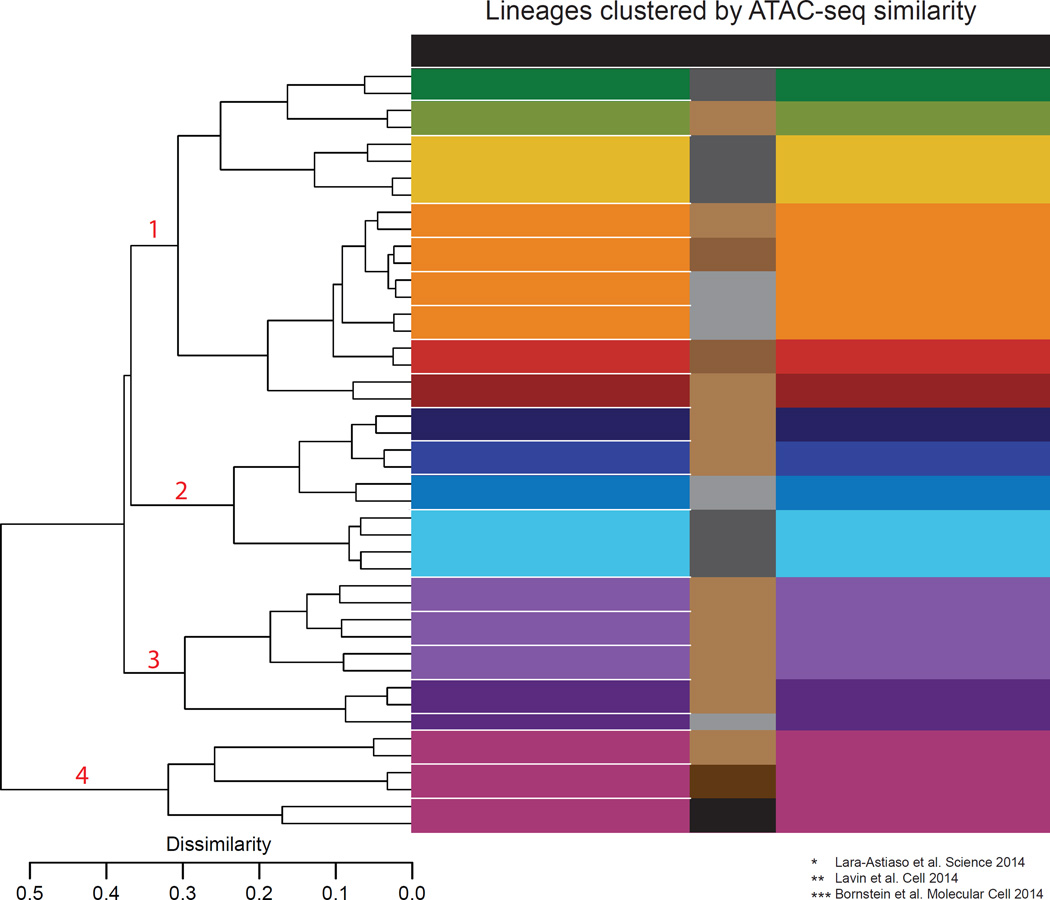
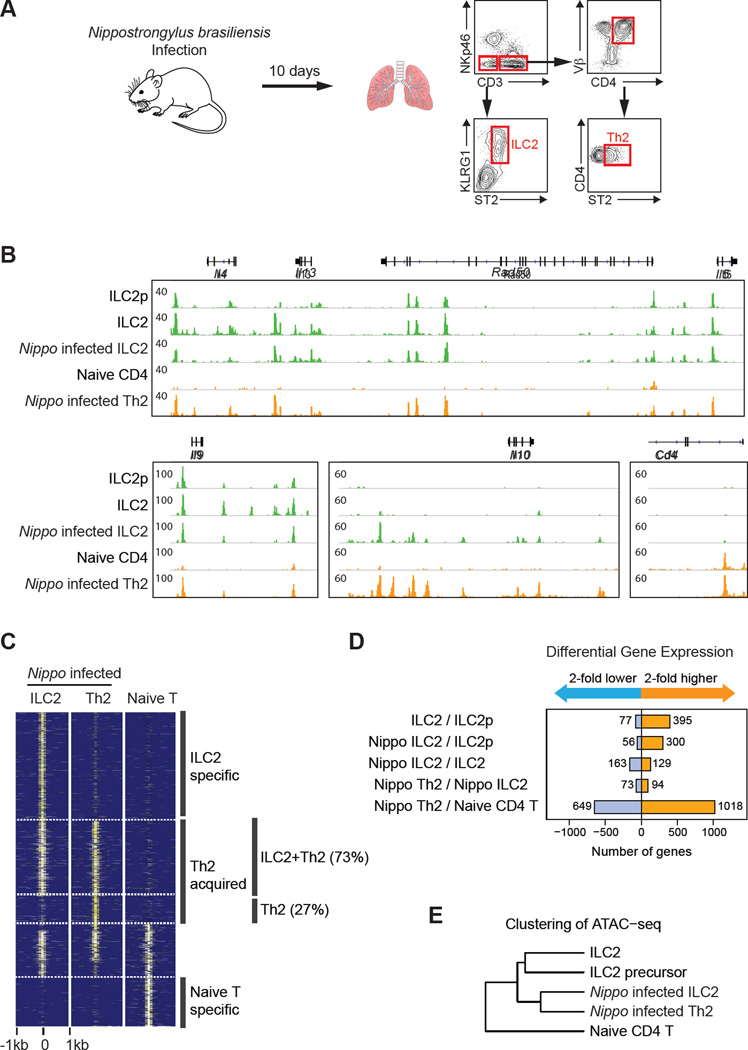
Innate lymphoid cells (ILCs) play key roles in host defense, barrier integrity, and homeostasis and mirror adaptive CD4(+) T helper (Th) cell subtypes in both usage of effector molecules and transcription factors. To better understand the relationship between ILC subsets and their Th cell counterparts, we measured genome-wide chromatin accessibility. We find that chromatin in proximity to effector genes is selectively accessible in ILCs prior to high-level transcription upon activation. Accessibility of these regions is acquired in a stepwise manner during development and changes little after in vitro or in vivo activation. Conversely, dramatic chromatin remodeling occurs in naive CD4(+) T cells during Th cell differentiation using a type-2-infection model. This alteration results in a substantial convergence of Th2 cells toward ILC2 regulomes. Our data indicate extensive sharing of regulatory circuitry across the innate and adaptive compartments of the immune system, in spite of their divergent developing pathways.
Published by Elsevier Inc.
Figures
References
-
- Artis D, Spits H. The biology of innate lymphoid cells. Nature. 2015;517:293–301. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
