

Proteolytic activation defines distinct lymphangiogenic mechanisms for VEGFC and VEGFD
- PMID: 27159393
- PMCID: PMC4887177
- DOI: 10.1172/JCI83967
Proteolytic activation defines distinct lymphangiogenic mechanisms for VEGFC and VEGFD
Abstract
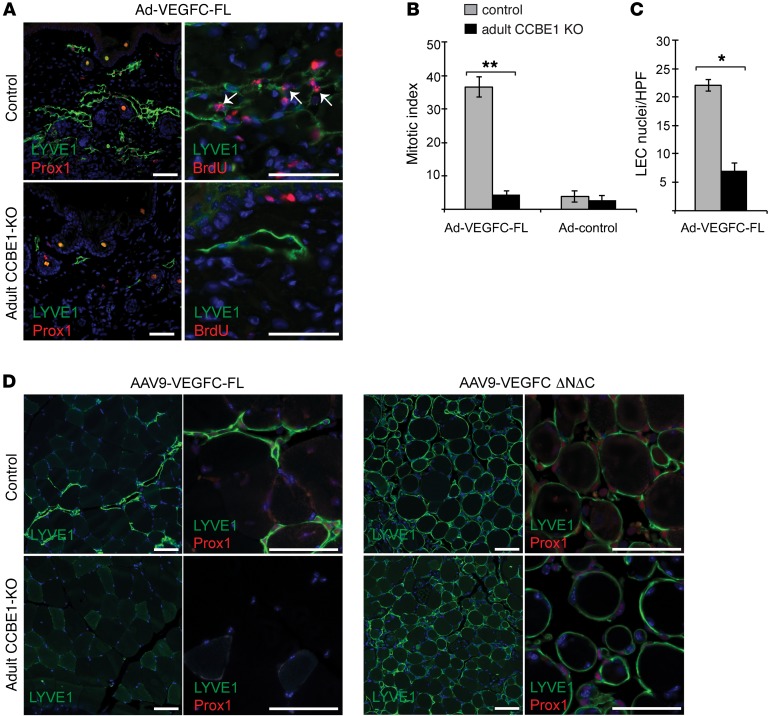
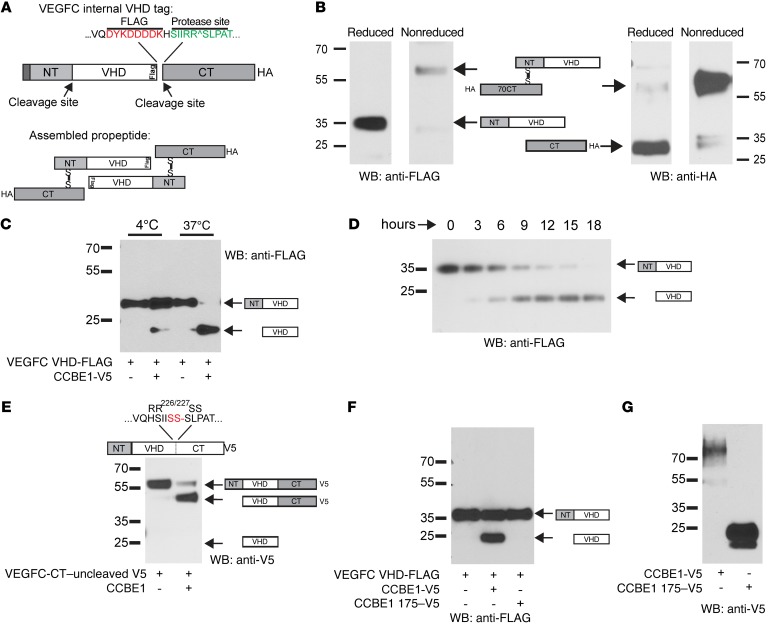
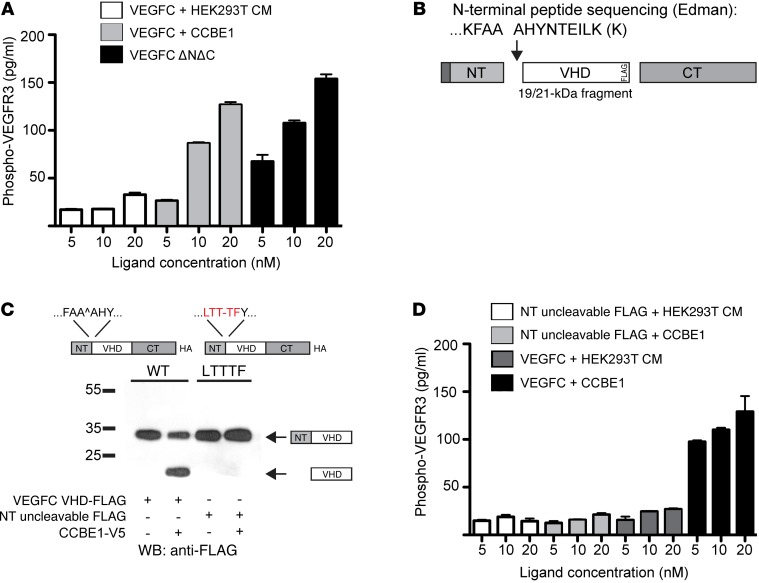
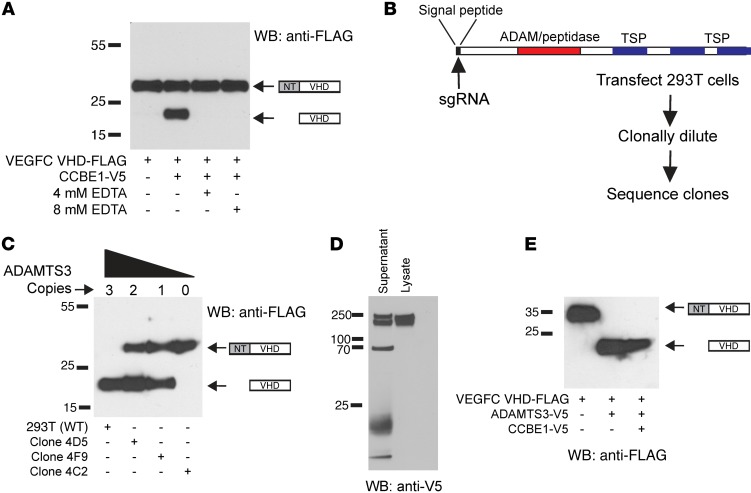
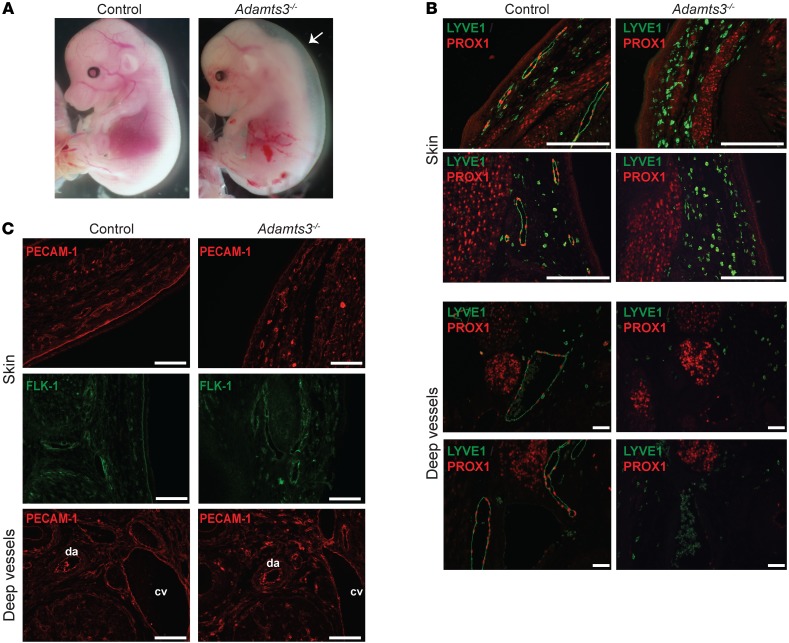
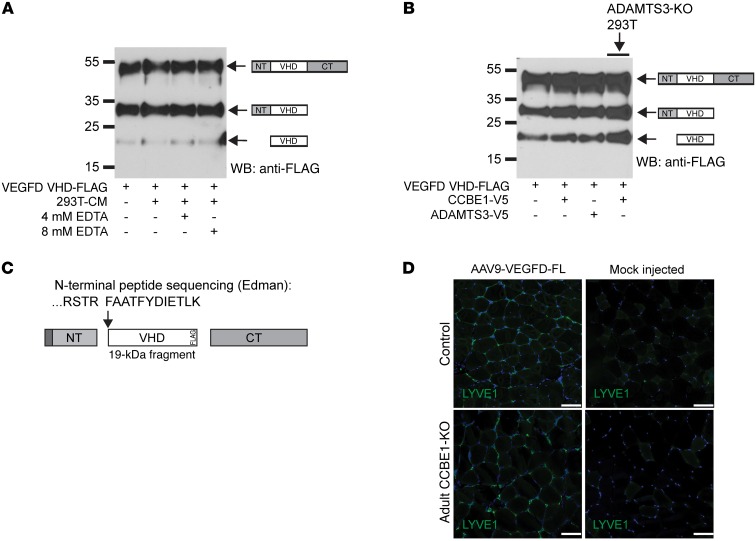
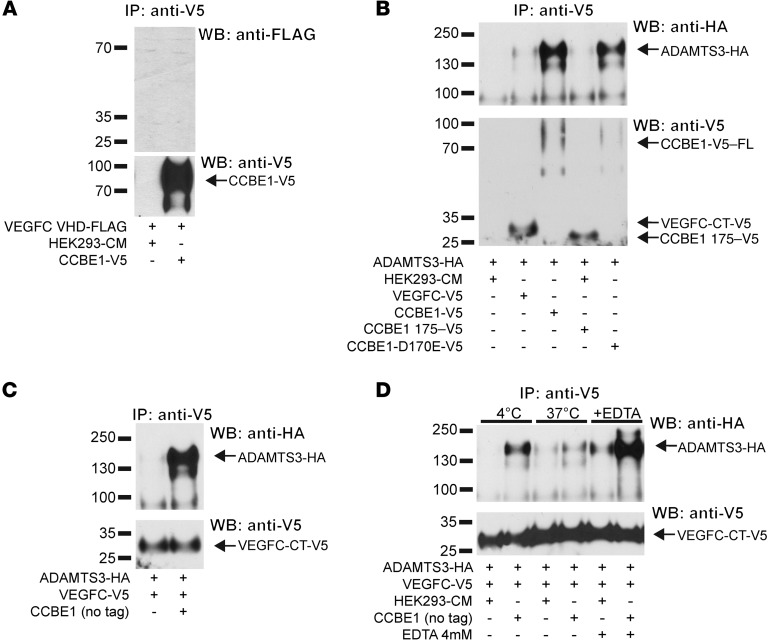
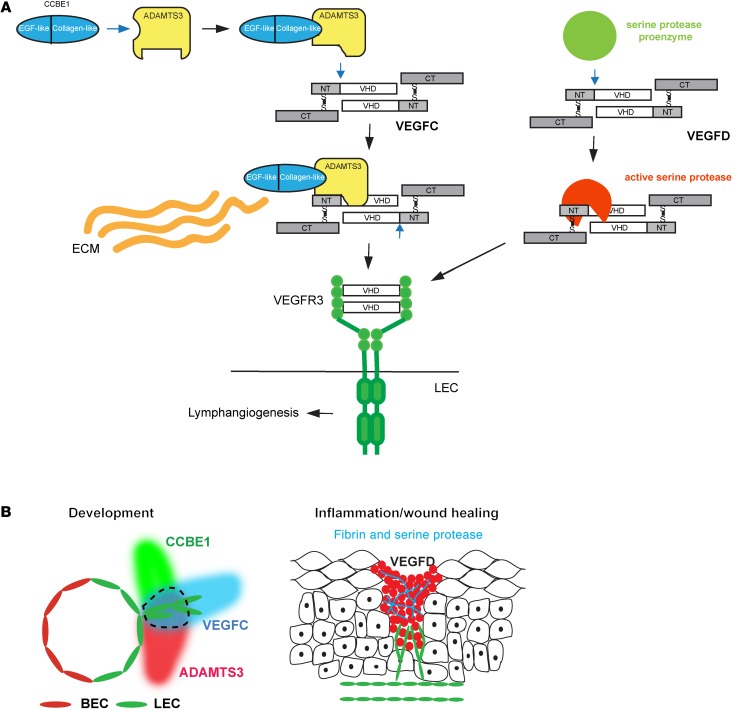
Lymphangiogenesis is supported by 2 homologous VEGFR3 ligands, VEGFC and VEGFD. VEGFC is required for lymphatic development, while VEGFD is not. VEGFC and VEGFD are proteolytically cleaved after cell secretion in vitro, and recent studies have implicated the protease a disintegrin and metalloproteinase with thrombospondin motifs 3 (ADAMTS3) and the secreted factor collagen and calcium binding EGF domains 1 (CCBE1) in this process. It is not well understood how ligand proteolysis is controlled at the molecular level or how this process regulates lymphangiogenesis, because these complex molecular interactions have been difficult to follow ex vivo and test in vivo. Here, we have developed and used biochemical and cellular tools to demonstrate that an ADAMTS3-CCBE1 complex can form independently of VEGFR3 and is required to convert VEGFC, but not VEGFD, into an active ligand. Consistent with these ex vivo findings, mouse genetic studies revealed that ADAMTS3 is required for lymphatic development in a manner that is identical to the requirement of VEGFC and CCBE1 for lymphatic development. Moreover, CCBE1 was required for in vivo lymphangiogenesis stimulated by VEGFC but not VEGFD. Together, these studies reveal that lymphangiogenesis is regulated by two distinct proteolytic mechanisms of ligand activation: one in which VEGFC activation by ADAMTS3 and CCBE1 spatially and temporally patterns developing lymphatics, and one in which VEGFD activation by a distinct proteolytic mechanism may be stimulated during inflammatory lymphatic growth.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
