

Metabolite concentrations, fluxes and free energies imply efficient enzyme usage
- PMID: 27159581
- PMCID: PMC4912430
- DOI: 10.1038/nchembio.2077
Metabolite concentrations, fluxes and free energies imply efficient enzyme usage
Abstract
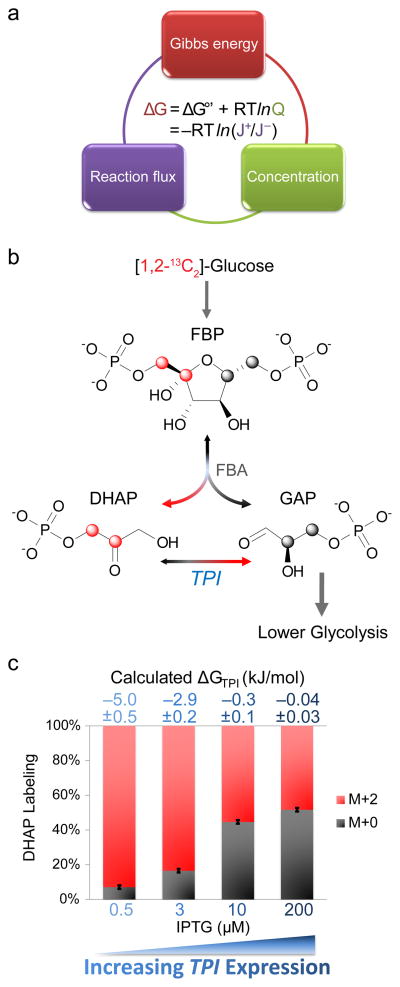
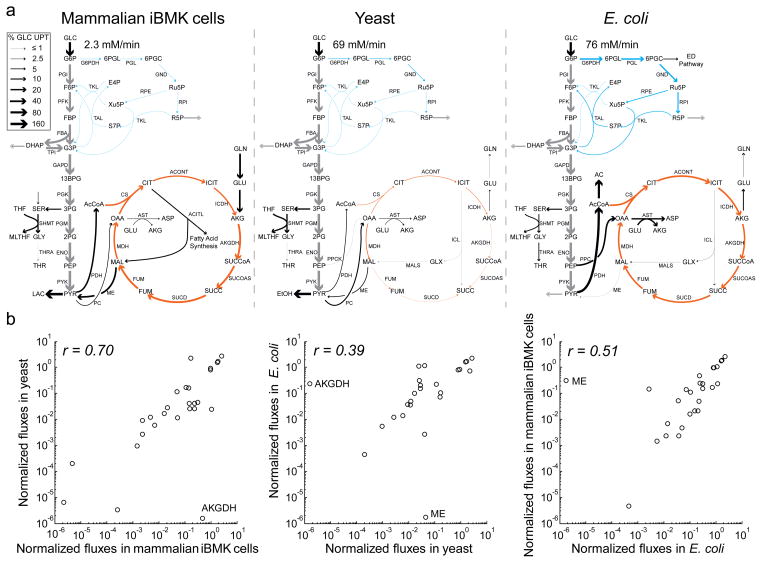
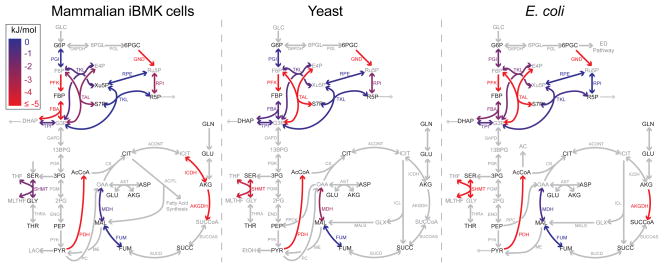
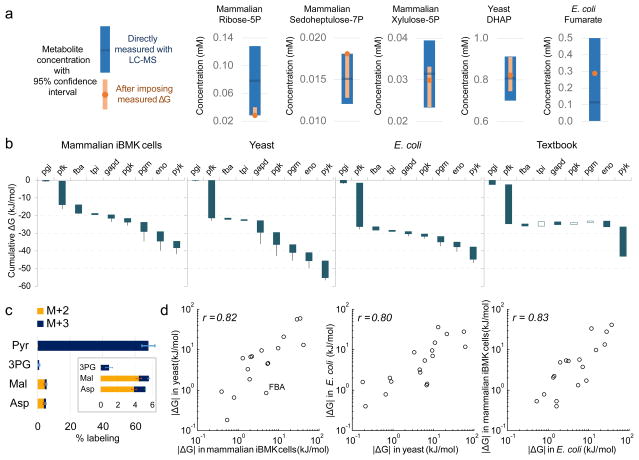
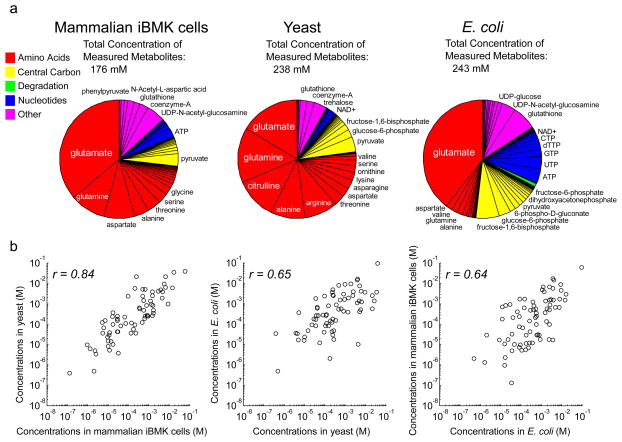
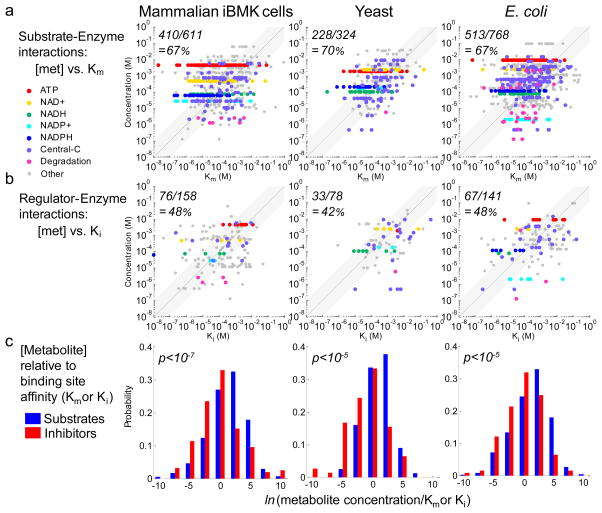
In metabolism, available free energy is limited and must be divided across pathway steps to maintain a negative ΔG throughout. For each reaction, ΔG is log proportional both to a concentration ratio (reaction quotient to equilibrium constant) and to a flux ratio (backward to forward flux). Here we use isotope labeling to measure absolute metabolite concentrations and fluxes in Escherichia coli, yeast and a mammalian cell line. We then integrate this information to obtain a unified set of concentrations and ΔG for each organism. In glycolysis, we find that free energy is partitioned so as to mitigate unproductive backward fluxes associated with ΔG near zero. Across metabolism, we observe that absolute metabolite concentrations and ΔG are substantially conserved and that most substrate (but not inhibitor) concentrations exceed the associated enzyme binding site dissociation constant (Km or Ki). The observed conservation of metabolite concentrations is consistent with an evolutionary drive to utilize enzymes efficiently given thermodynamic and osmotic constraints.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Joshi A, Palsson BO. Metabolic dynamics in the human red cell. Part III—Metabolic reaction rates. Journal of Theoretical Biology. 1990;142:41–68. - PubMed
-
- Chassagnole C, Noisommit-Rizzi N, Schmid JW, Mauch K, Reuss M. Dynamic modeling of the central carbon metabolism of Escherichia coli. Biotechnol Bioeng. 2002;79:53–73. - PubMed
-
- van Eunen K, et al. Measuring enzyme activities under standardized in vivo-like conditions for systems biology. FEBS J. 2010;277:749–760. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
