

The Tumor Suppressor Hace1 Is a Critical Regulator of TNFR1-Mediated Cell Fate
- PMID: 27160902
- PMCID: PMC4893156
- DOI: 10.1016/j.celrep.2016.04.032
The Tumor Suppressor Hace1 Is a Critical Regulator of TNFR1-Mediated Cell Fate
Erratum in
-
The Tumor Suppressor Hace1 Is a Critical Regulator of TNFR1-Mediated Cell Fate.Cell Rep. 2016 Sep 20;16(12):3414. doi: 10.1016/j.celrep.2016.08.072. Epub 2016 Sep 20. Cell Rep. 2016. PMID: 27653700 Free PMC article. No abstract available.
Abstract
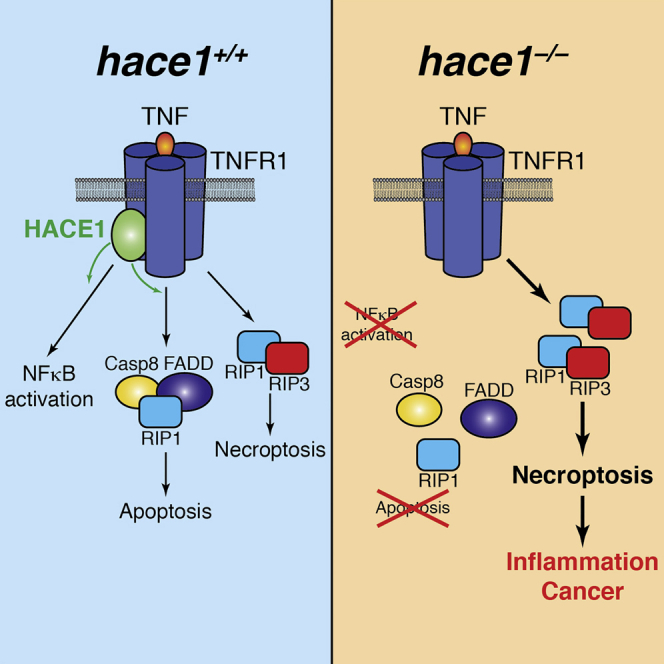
The HECT domain E3 ligase HACE1 has been identified as a tumor suppressor in multiple cancers. Here, we report that HACE1 is a central gatekeeper of TNFR1-induced cell fate. Genetic inactivation of HACE1 inhibits TNF-stimulated NF-κB activation and TNFR1-NF-κB-dependent pathogen clearance in vivo. Moreover, TNF-induced apoptosis was impaired in hace1 mutant cells and knockout mice in vivo. Mechanistically, HACE1 is essential for the ubiquitylation of the adaptor protein TRAF2 and formation of the apoptotic caspase-8 effector complex. Intriguingly, loss of HACE1 does not impair TNFR1-mediated necroptotic cell fate via RIP1 and RIP3 kinases. Loss of HACE1 predisposes animals to colonic inflammation and carcinogenesis in vivo, which is markedly alleviated by genetic inactivation of RIP3 kinase and TNFR1. Thus, HACE1 controls TNF-elicited cell fate decisions and exerts tumor suppressor and anti-inflammatory activities via a TNFR1-RIP3 kinase-necroptosis pathway.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Figures







Comment in
-
TNF-driven cell fate: till HACE do us part.Oncotarget. 2016 Jul 19;7(29):44871-44872. doi: 10.18632/oncotarget.10168. Oncotarget. 2016. PMID: 27343549 Free PMC article. No abstract available.
References
-
- Anglesio M.S., Evdokimova V., Melnyk N., Zhang L., Fernandez C.V., Grundy P.E., Leach S., Marra M.A., Brooks-Wilson A.R., Penninger J., Sorensen P.H. Differential expression of a novel ankyrin containing E3 ubiquitin-protein ligase, Hace1, in sporadic Wilms’ tumor versus normal kidney. Hum. Mol. Genet. 2004;13:2061–2074. - PubMed
-
- Beg A.A., Baltimore D. An essential role for NF-kappaB in preventing TNF-alpha-induced cell death. Science. 1996;274:782–784. - PubMed
-
- Bergink S., Jentsch S. Principles of ubiquitin and SUMO modifications in DNA repair. Nature. 2009;458:461–467. - PubMed
-
- Bernassola F., Ciechanover A., Melino G. The ubiquitin proteasome system and its involvement in cell death pathways. Cell Death Differ. 2010;17:1–3. - PubMed
-
- Bhoj V.G., Chen Z.J. Ubiquitylation in innate and adaptive immunity. Nature. 2009;458:430–437. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
