

Myelinating satellite oligodendrocytes are integrated in a glial syncytium constraining neuronal high-frequency activity
- PMID: 27161034
- PMCID: PMC4866043
- DOI: 10.1038/ncomms11298
Myelinating satellite oligodendrocytes are integrated in a glial syncytium constraining neuronal high-frequency activity
Abstract
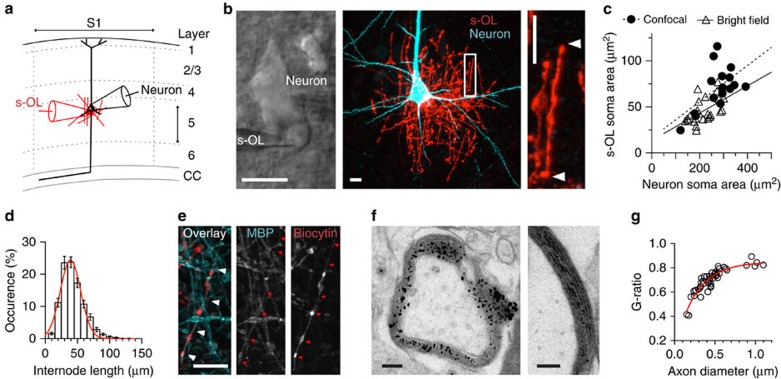
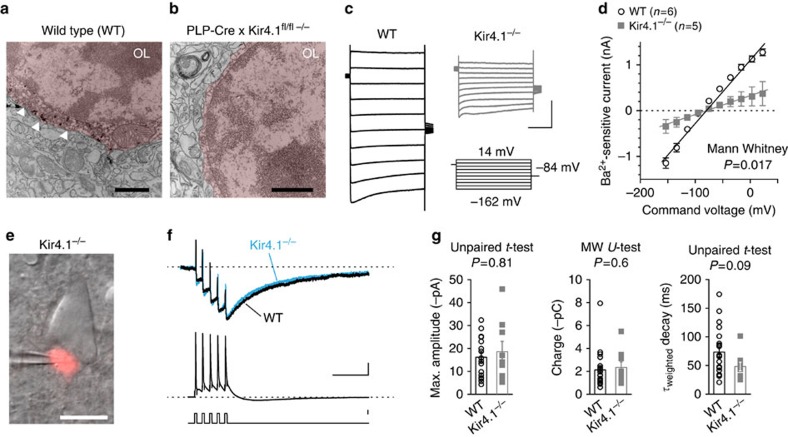
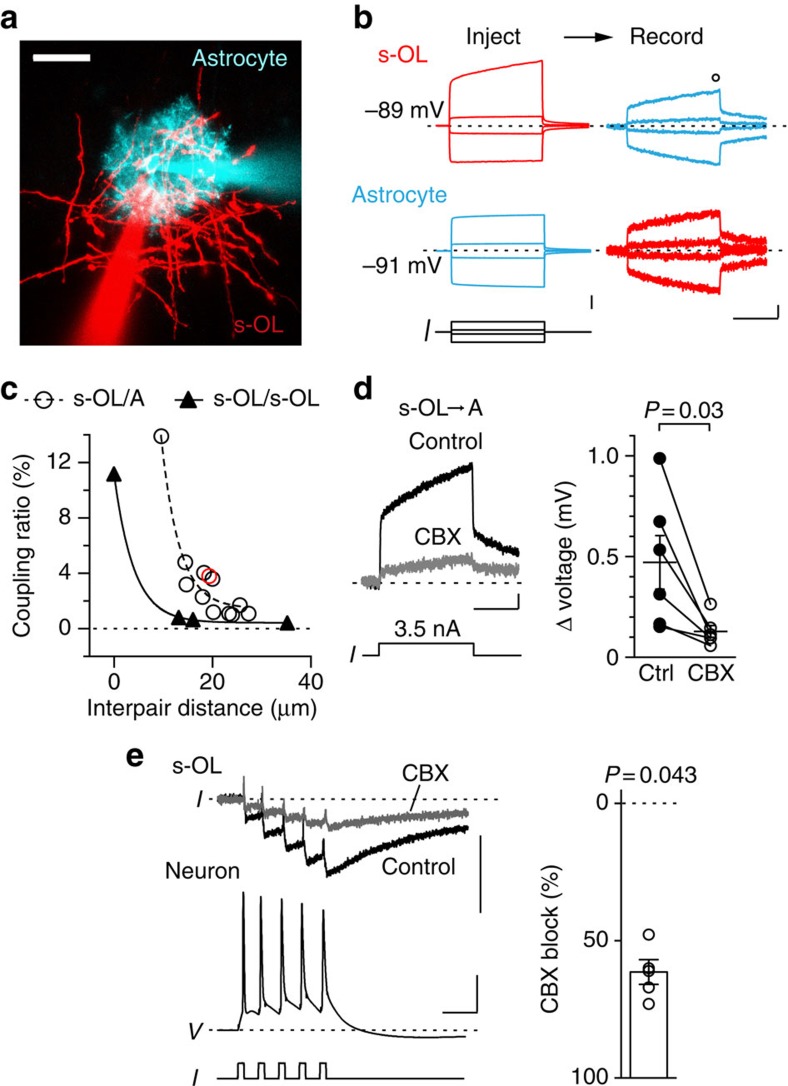
Satellite oligodendrocytes (s-OLs) are closely apposed to the soma of neocortical layer 5 pyramidal neurons but their properties and functional roles remain unresolved. Here we show that s-OLs form compact myelin and action potentials of the host neuron evoke precisely timed Ba(2+)-sensitive K(+) inward rectifying (Kir) currents in the s-OL. Unexpectedly, the glial K(+) inward current does not require oligodendrocytic Kir4.1. Action potential-evoked Kir currents are in part mediated by gap-junction coupling with neighbouring OLs and astrocytes that form a syncytium around the pyramidal cell body. Computational modelling predicts that glial Kir constrains the perisomatic [K(+)]o increase most importantly during high-frequency action potentials. Consistent with these predictions neurons with s-OLs showed a reduced probability for action potential burst firing during [K(+)]o elevations. These data suggest that s-OLs are integrated into a glial syncytium for the millisecond rapid K(+) uptake limiting activity-dependent [K(+)]o increase in the perisomatic neuron domain.
Figures





References
-
- Min R. & Nevian T. Astrocyte signaling controls spike timing-dependent depression at neocortical synapses. Nat. Neurosci. 15, 746–753 (2012). - PubMed
-
- Chen J. et al. Heterosynaptic long-term depression mediated by ATP released from astrocytes. Glia 61, 178–191 (2013). - PubMed
-
- Morquette P. et al. An astrocyte-dependent mechanism for neuronal rhythmogenesis. Nat. Neurosci. 18, 844–854 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
