

Structural Basis of Diverse Homophilic Recognition by Clustered α- and β-Protocadherins
- PMID: 27161523
- PMCID: PMC4873334
- DOI: 10.1016/j.neuron.2016.04.004
Structural Basis of Diverse Homophilic Recognition by Clustered α- and β-Protocadherins
Abstract
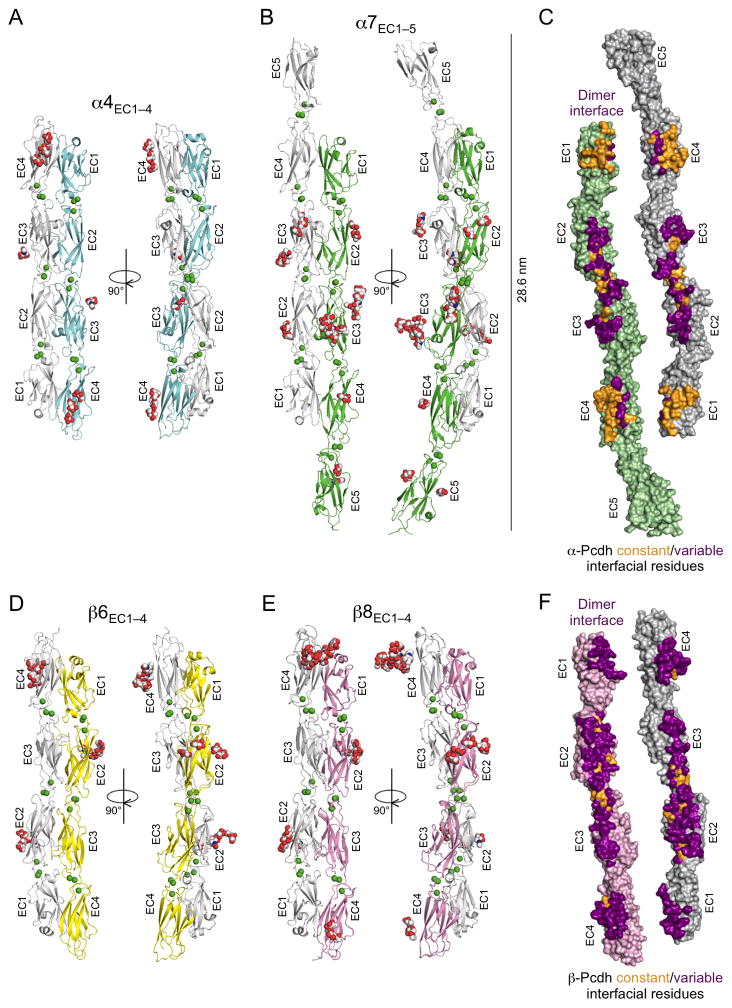
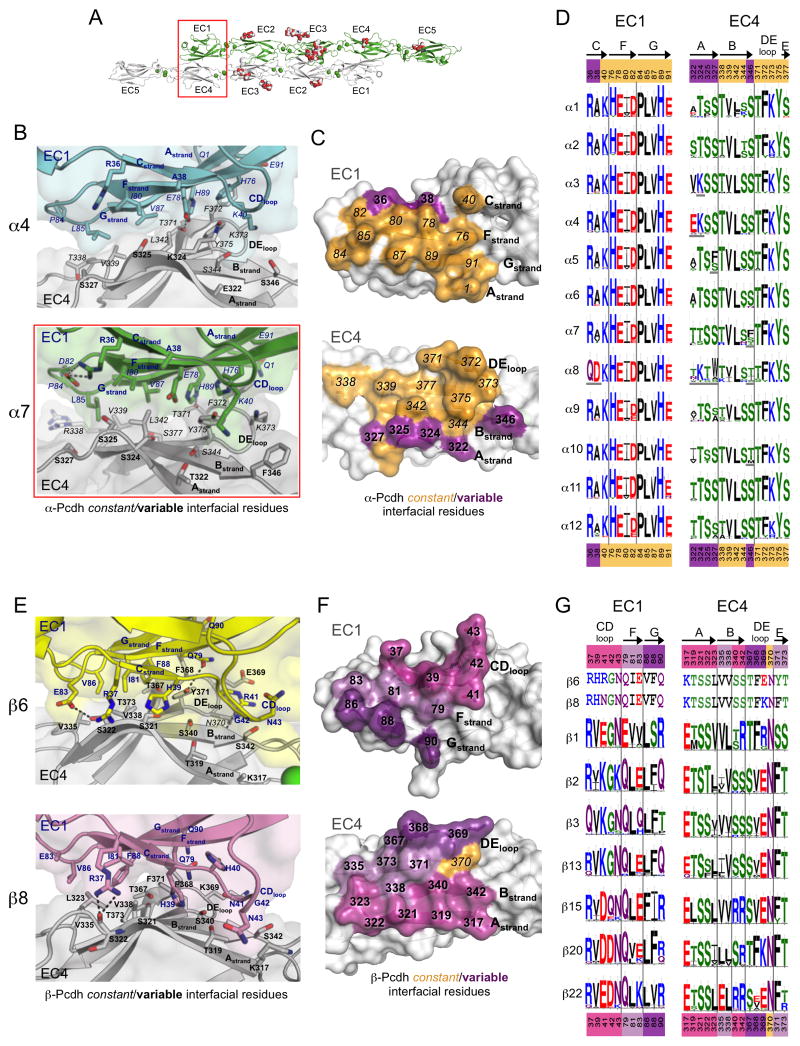
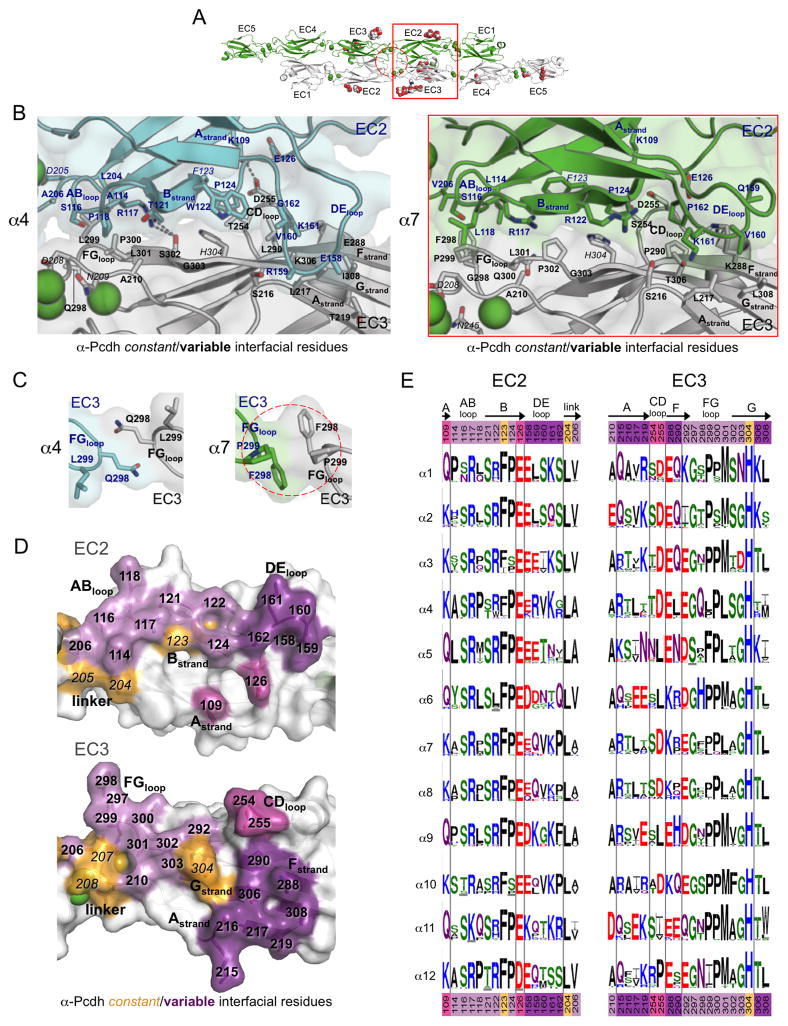
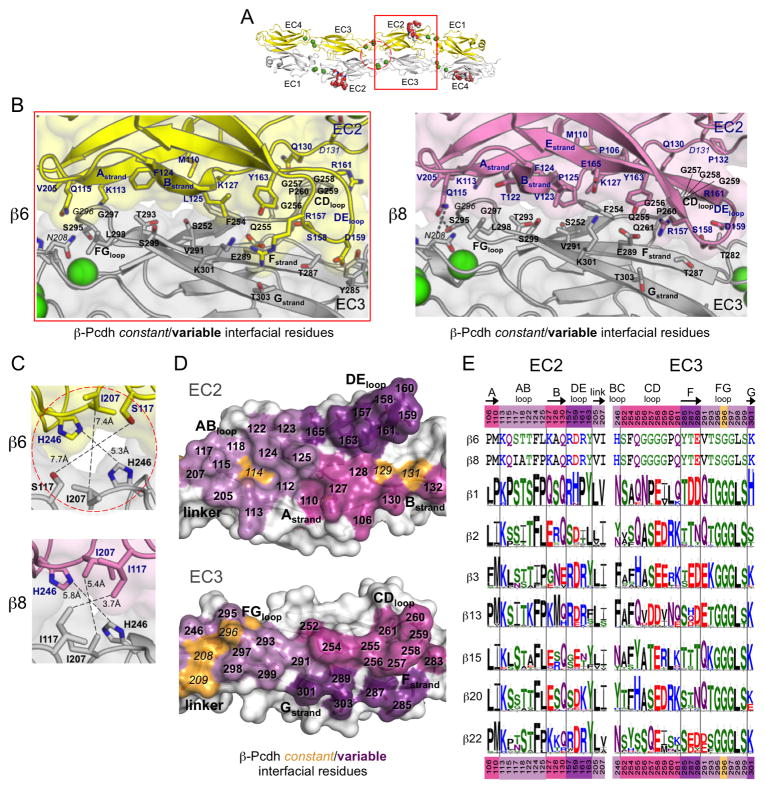
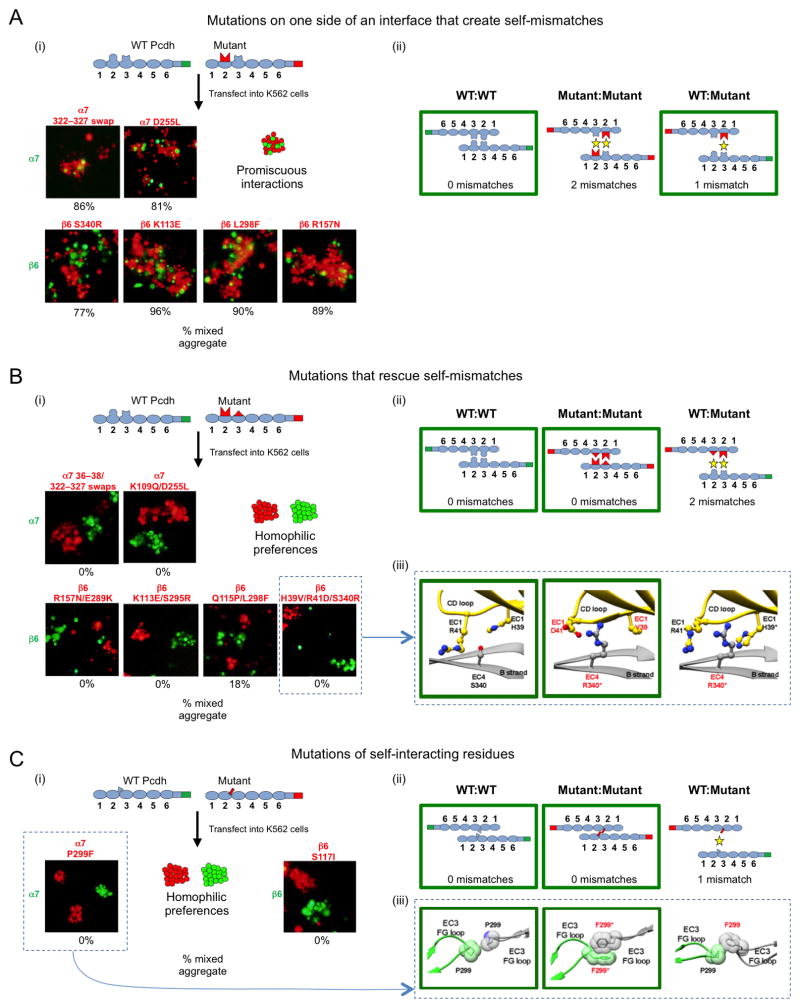
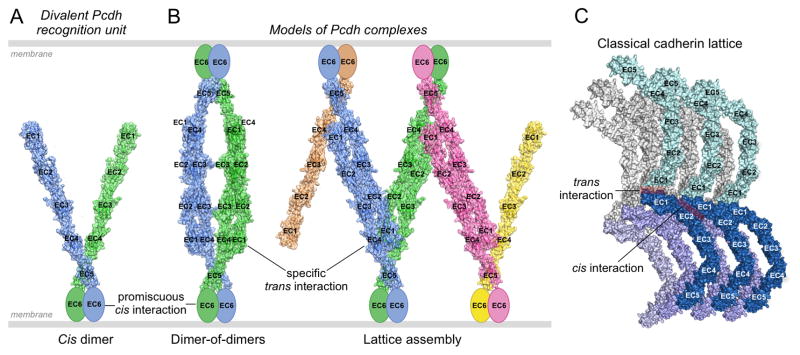
Clustered protocadherin proteins (α-, β-, and γ-Pcdhs) provide a high level of cell-surface diversity to individual vertebrate neurons, engaging in highly specific homophilic interactions to mediate important roles in mammalian neural circuit development. How Pcdhs bind homophilically through their extracellular cadherin (EC) domains among dozens of highly similar isoforms has not been determined. Here, we report crystal structures for extracellular regions from four mouse Pcdh isoforms (α4, α7, β6, and β8), revealing a canonical head-to-tail interaction mode for homophilic trans dimers comprising primary intermolecular EC1:EC4 and EC2:EC3 interactions. A subset of trans interface residues exhibit isoform-specific conservation, suggesting roles in recognition specificity. Mutation of these residues, along with trans-interacting partner residues, altered the specificities of Pcdh interactions. Together, these data show how sequence variation among Pcdh isoforms encodes their diverse strict homophilic recognition specificities, which are required for their key roles in neural circuit assembly.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Aricescu AR, Jones EY. Immunoglobulin superfamily cell adhesion molecules: zippers and signals. Curr Opin Cell Biol. 2007;19:543–50. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
