

Cos-Seq for high-throughput identification of drug target and resistance mechanisms in the protozoan parasite Leishmania
- PMID: 27162331
- PMCID: PMC4889358
- DOI: 10.1073/pnas.1520693113
Cos-Seq for high-throughput identification of drug target and resistance mechanisms in the protozoan parasite Leishmania
Abstract
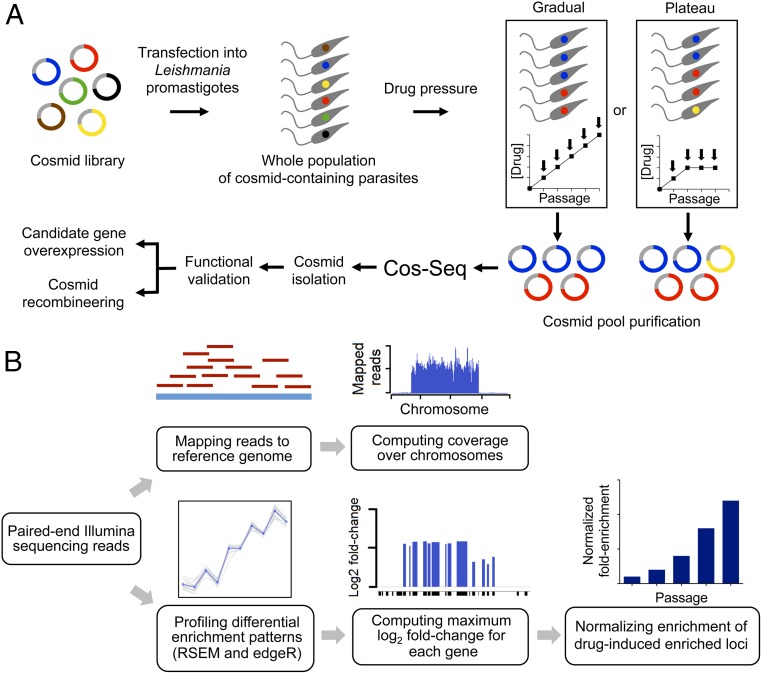
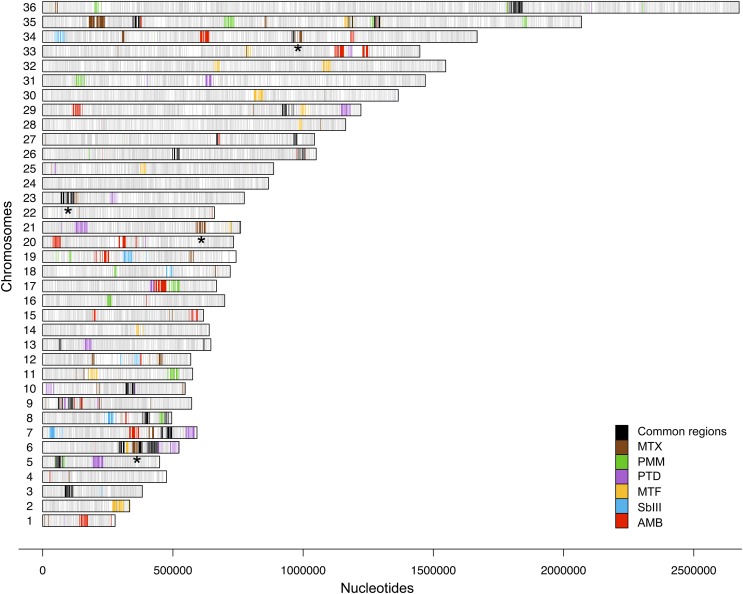
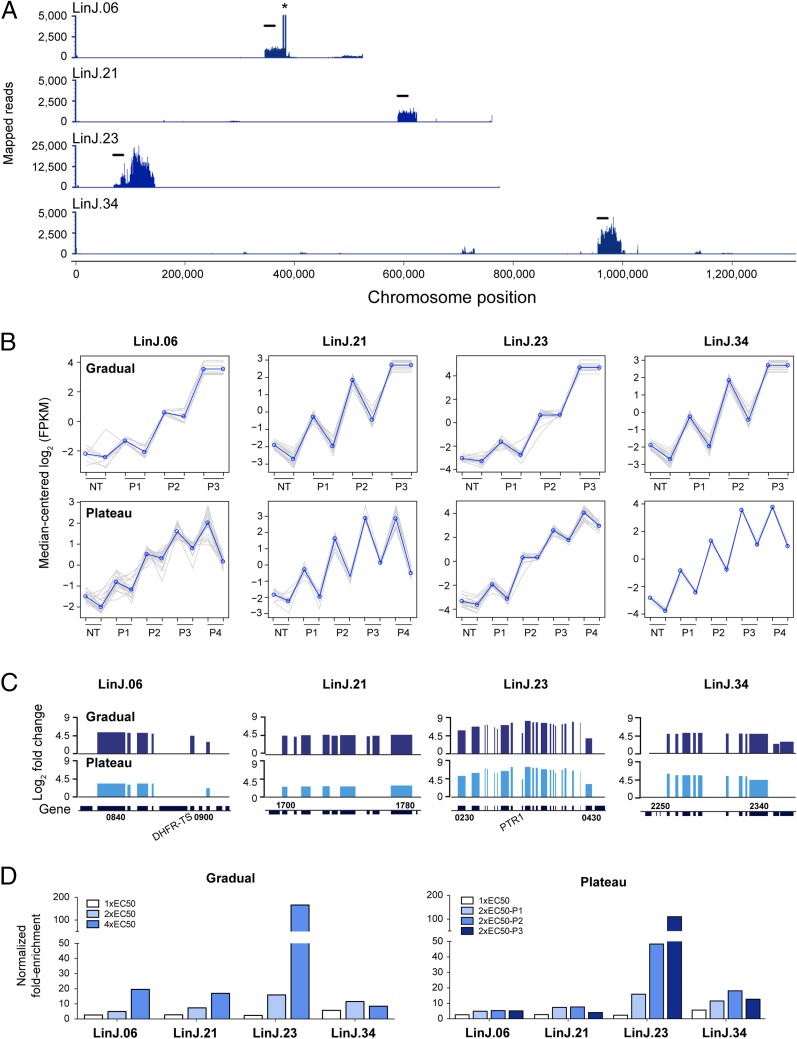
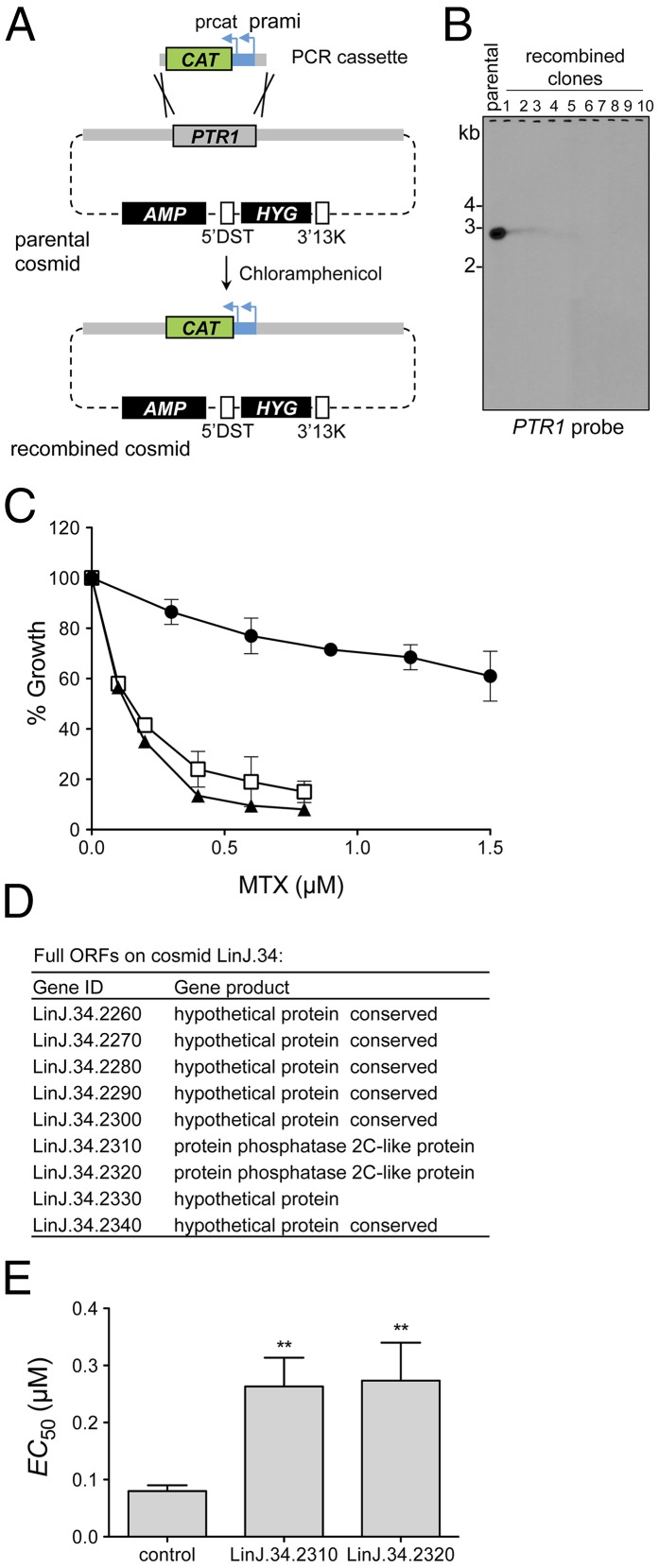
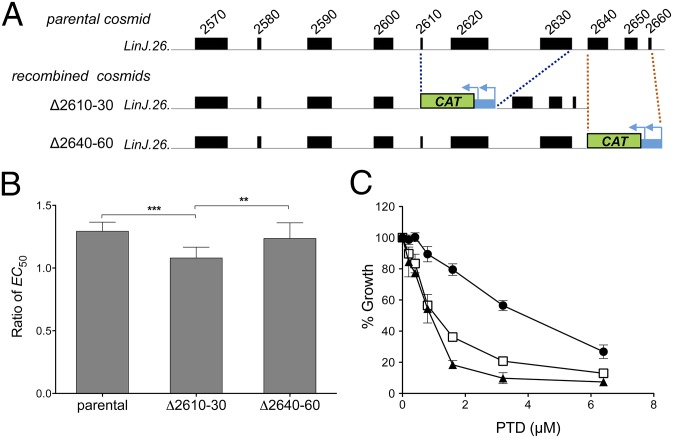
Innovative strategies are needed to accelerate the identification of antimicrobial drug targets and resistance mechanisms. Here we develop a sensitive method, which we term Cosmid Sequencing (or "Cos-Seq"), based on functional cloning coupled to next-generation sequencing. Cos-Seq identified >60 loci in the Leishmania genome that were enriched via drug selection with methotrexate and five major antileishmanials (antimony, miltefosine, paromomycin, amphotericin B, and pentamidine). Functional validation highlighted both known and previously unidentified drug targets and resistance genes, including novel roles for phosphatases in resistance to methotrexate and antimony, for ergosterol and phospholipid metabolism genes in resistance to miltefosine, and for hypothetical proteins in resistance to paromomycin, amphothericin B, and pentamidine. Several genes/loci were also found to confer resistance to two or more antileishmanials. This screening method will expedite the discovery of drug targets and resistance mechanisms and is easily adaptable to other microorganisms.
Keywords: Cos-Seq; Leishmania; functional cloning; next-generation sequencing; resistance.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
High-throughput Cos-Seq screen with intracellular Leishmania infantum for the discovery of novel drug-resistance mechanisms.Int J Parasitol Drugs Drug Resist. 2018 Aug;8(2):165-173. doi: 10.1016/j.ijpddr.2018.03.004. Epub 2018 Mar 16. Int J Parasitol Drugs Drug Resist. 2018. PMID: 29602064 Free PMC article.
-
CRISPR-Cas9 high-throughput screening to study drug resistance in Leishmania infantum.mBio. 2024 Jul 17;15(7):e0047724. doi: 10.1128/mbio.00477-24. Epub 2024 Jun 12. mBio. 2024. PMID: 38864609 Free PMC article.
-
Cos-Seq: A High-Throughput Gain-of-Function Screen for Drug Resistance Studies in Leishmania.Methods Mol Biol. 2019;1971:141-167. doi: 10.1007/978-1-4939-9210-2_7. Methods Mol Biol. 2019. PMID: 30980302
-
Functional cloning as a means to identify Leishmania genes involved in drug resistance.Mini Rev Med Chem. 2006 Feb;6(2):123-9. doi: 10.2174/138955706775476028. Mini Rev Med Chem. 2006. PMID: 16472181 Review.
-
Drug resistance analysis by next generation sequencing in Leishmania.Int J Parasitol Drugs Drug Resist. 2014 Sep 22;5(1):26-35. doi: 10.1016/j.ijpddr.2014.09.005. eCollection 2015 Apr. Int J Parasitol Drugs Drug Resist. 2014. PMID: 25941624 Free PMC article. Review.
Cited by
-
Of Drugs and Trypanosomatids: New Tools and Knowledge to Reduce Bottlenecks in Drug Discovery.Genes (Basel). 2020 Jun 29;11(7):722. doi: 10.3390/genes11070722. Genes (Basel). 2020. PMID: 32610603 Free PMC article. Review.
-
Omics Approaches in Drug Development against Leishmaniasis: Current Scenario and Future Prospects.Pathogens. 2022 Dec 26;12(1):39. doi: 10.3390/pathogens12010039. Pathogens. 2022. PMID: 36678387 Free PMC article. Review.
-
Identification of Resistance Determinants for a Promising Antileishmanial Oxaborole Series.Microorganisms. 2021 Jun 29;9(7):1408. doi: 10.3390/microorganisms9071408. Microorganisms. 2021. PMID: 34210040 Free PMC article.
-
Antimony resistance and gene expression in Leishmania: spotlight on molecular and proteomic aspects.Parasitology. 2024 Jan;151(1):1-14. doi: 10.1017/S0031182023001129. Epub 2023 Nov 28. Parasitology. 2024. PMID: 38012864 Free PMC article. Review.
-
Potential selection of antimony and methotrexate cross-resistance in Leishmania infantum circulating strains.PLoS Negl Trop Dis. 2024 Feb 29;18(2):e0012015. doi: 10.1371/journal.pntd.0012015. eCollection 2024 Feb. PLoS Negl Trop Dis. 2024. PMID: 38422164 Free PMC article.
References
-
- World, Health Organization . Sustaining the Drive to Overcome the Global Impact of Neglected Tropical Diseases: Second WHO Report on Neglected Tropical Diseases. World Health Organization; Geneva, Switzerland: 2013.
-
- Ouellette M, Drummelsmith J, Papadopoulou B. Leishmaniasis: Drugs in the clinic, resistance and new developments. Drug Resist Update. 2004;7(4–5):257–266. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
