

GPCRs Direct Germline Development and Somatic Gonad Function in Planarians
- PMID: 27163480
- PMCID: PMC4862687
- DOI: 10.1371/journal.pbio.1002457
GPCRs Direct Germline Development and Somatic Gonad Function in Planarians
Abstract
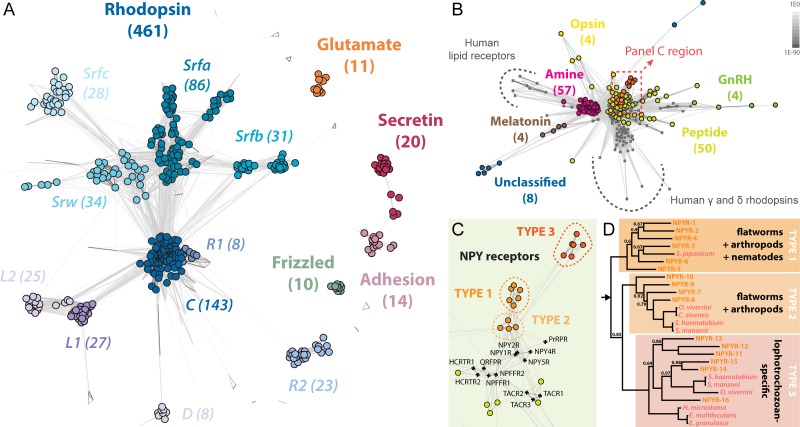
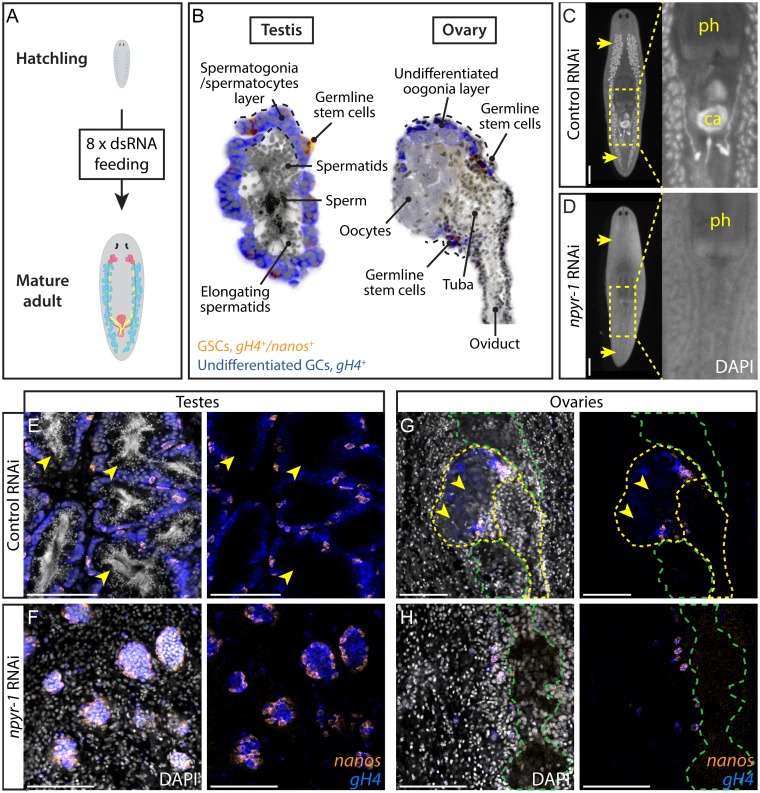
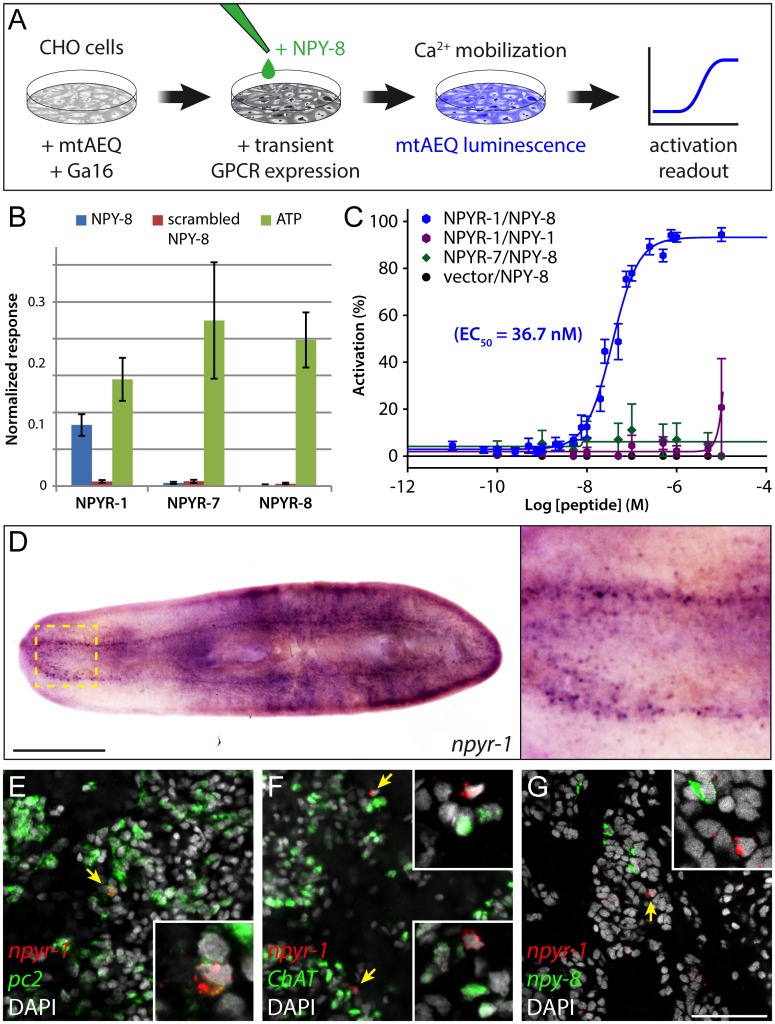
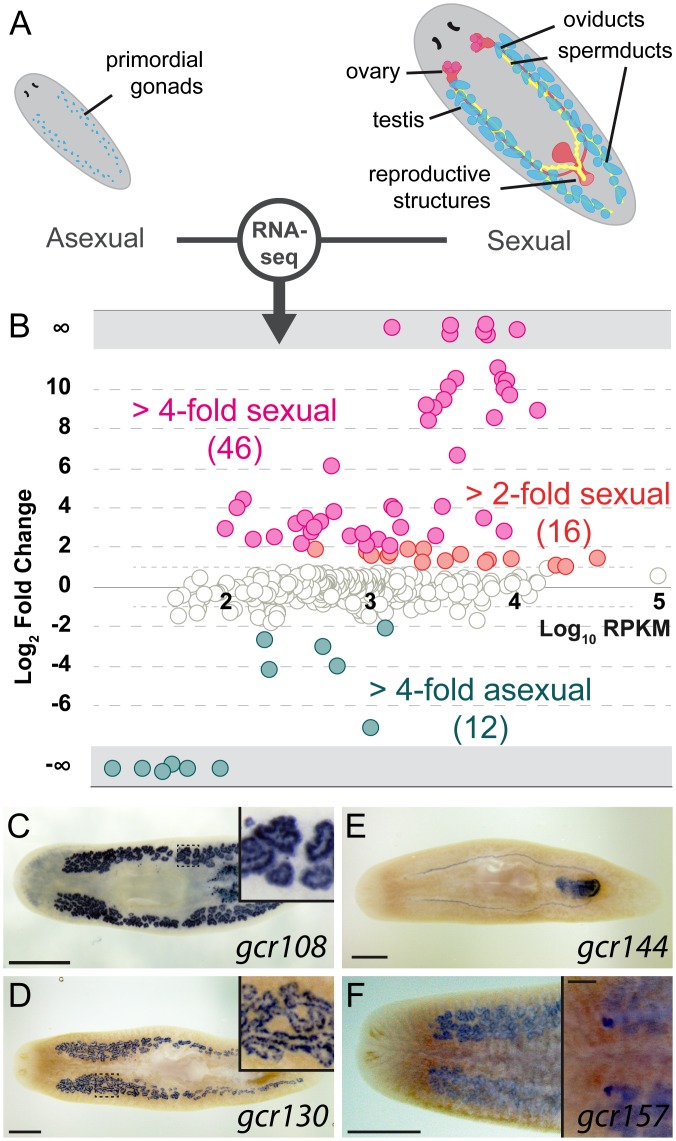
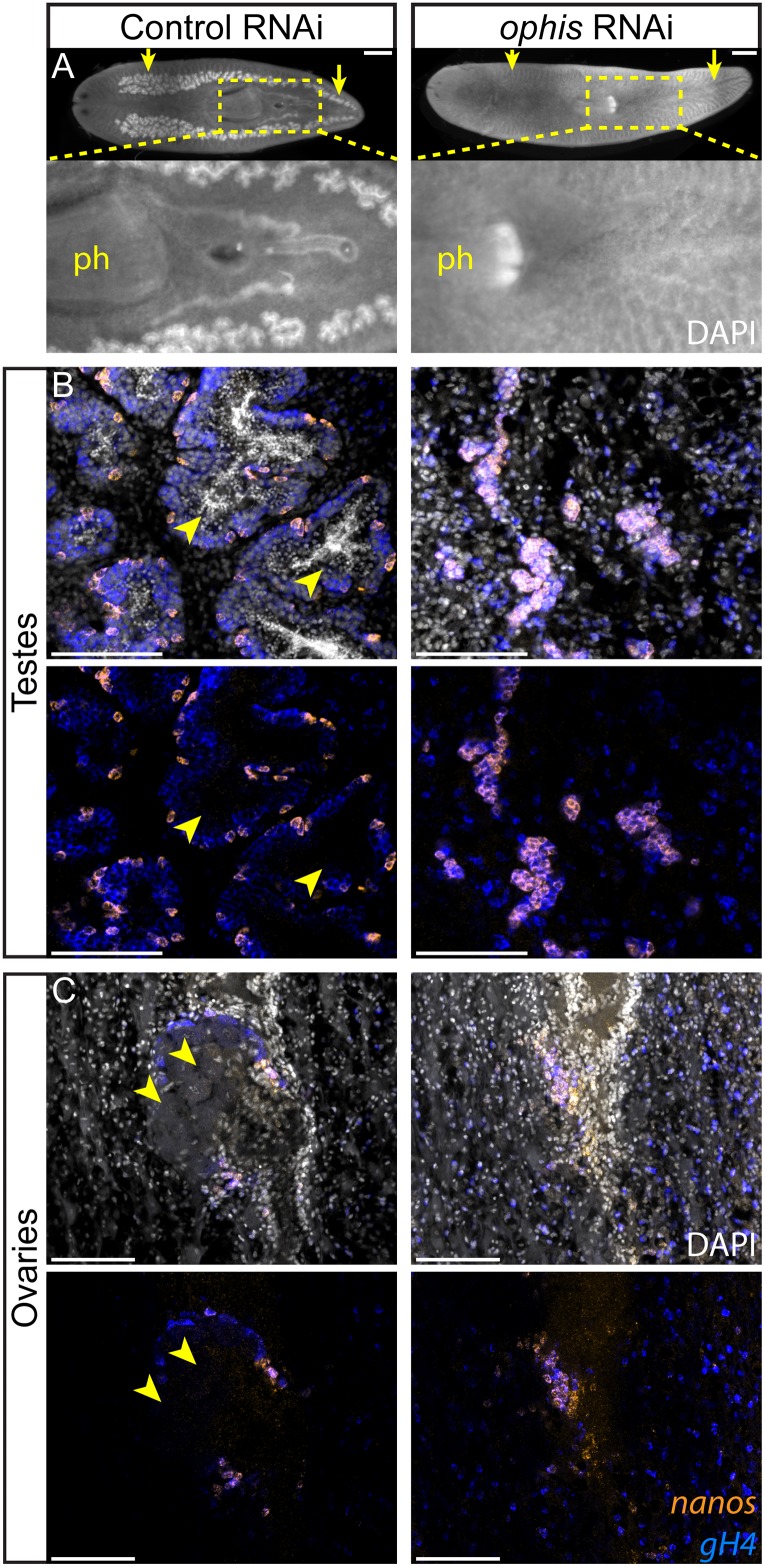
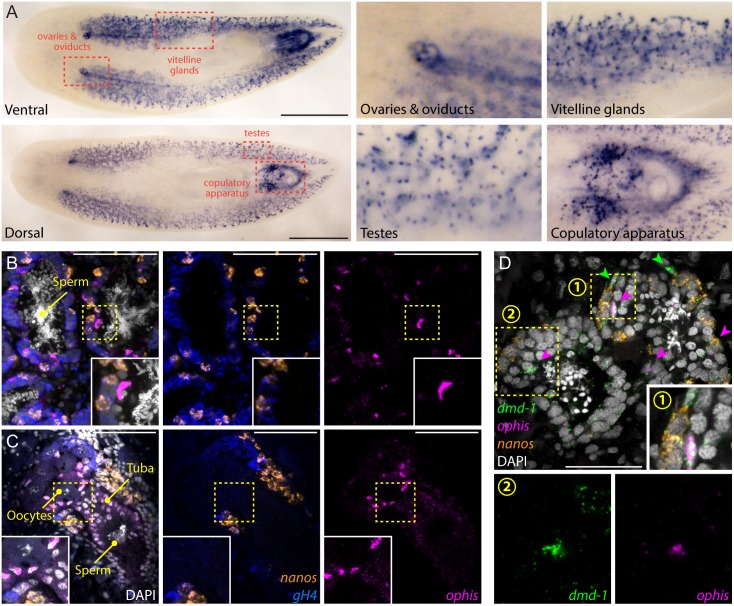
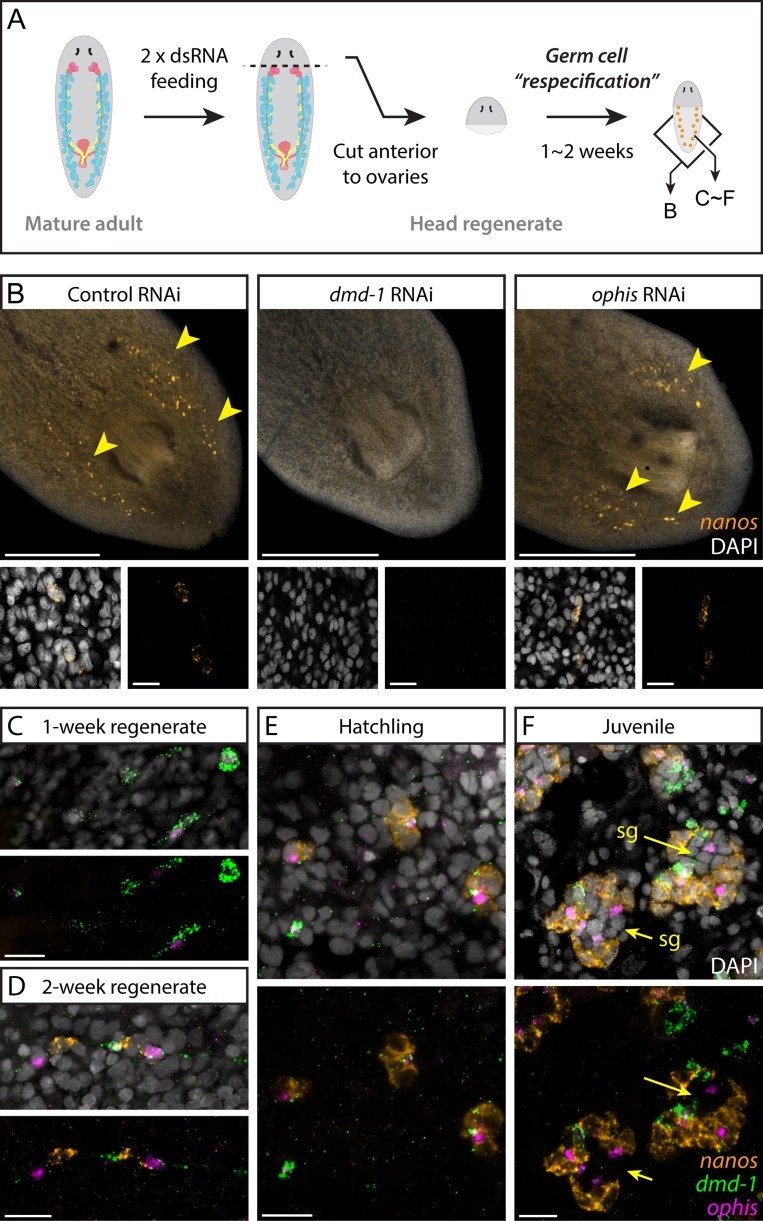
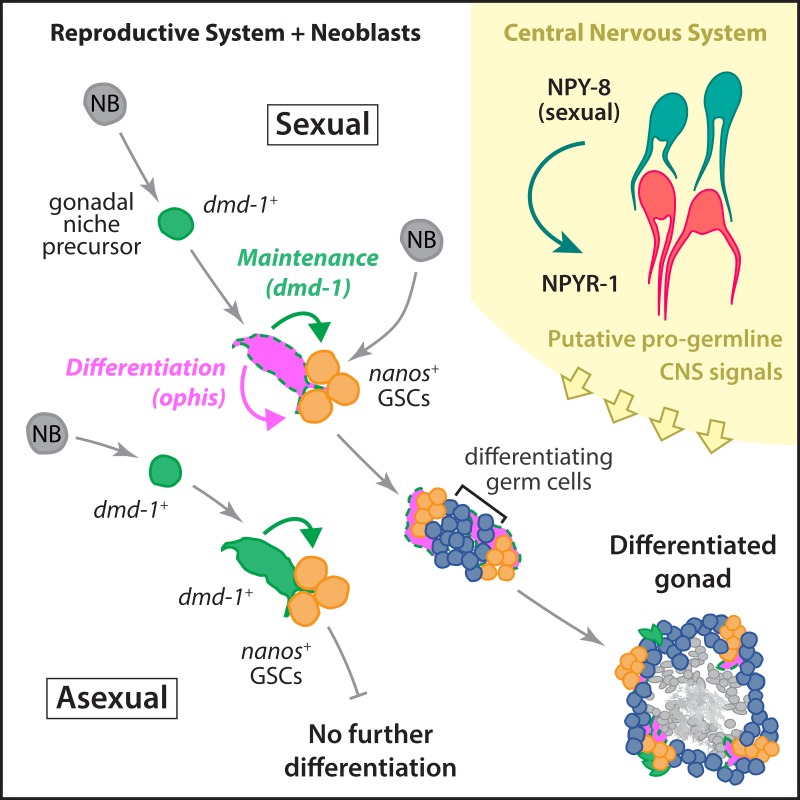
Planarians display remarkable plasticity in maintenance of their germline, with the ability to develop or dismantle reproductive tissues in response to systemic and environmental cues. Here, we investigated the role of G protein-coupled receptors (GPCRs) in this dynamic germline regulation. By genome-enabled receptor mining, we identified 566 putative planarian GPCRs and classified them into conserved and phylum-specific subfamilies. We performed a functional screen to identify NPYR-1 as the cognate receptor for NPY-8, a neuropeptide required for sexual maturation and germ cell differentiation. Similar to NPY-8, knockdown of this receptor results in loss of differentiated germ cells and sexual maturity. NPYR-1 is expressed in neuroendocrine cells of the central nervous system and can be activated specifically by NPY-8 in cell-based assays. Additionally, we screened the complement of GPCRs with expression enriched in sexually reproducing planarians, and identified an orphan chemoreceptor family member, ophis, that controls differentiation of germline stem cells (GSCs). ophis is expressed in somatic cells of male and female gonads, as well as in accessory reproductive tissues. We have previously shown that somatic gonadal cells are required for male GSC specification and maintenance in planarians. However, ophis is not essential for GSC specification or maintenance and, therefore, defines a secondary role for planarian gonadal niche cells in promoting GSC differentiation. Our studies uncover the complement of planarian GPCRs and reveal previously unappreciated roles for these receptors in systemic and local (i.e., niche) regulation of germ cell development.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Tsutsumi M, Zhou W, Millar RP, Mellon PL, Roberts JL, Flanagan CA, et al. Cloning and functional expression of a mouse gonadotropin-releasing hormone receptor. Mol Endocrinol. 1992;6: 1163–1169. - PubMed
-
- Millar RP, Lu ZL, Pawson AJ, Flanagan CA, Morgan K, Maudsley SR. Gonadotropin-releasing hormone receptors. Endocr Rev. 2004;25: 235–275. - PubMed
-
- Cole LW, Sidis Y, Zhang C, Quinton R, Plummer L, Pignatelli D, et al. Mutations in prokineticin 2 and prokineticin receptor 2 genes in human gonadotrophin-releasing hormone deficiency: molecular genetics and clinical spectrum. J Clin Endocrinol Metab. 2008;93: 3551–3559. 10.1210/jc.2007-2654 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
