

Extracellular acidification stimulates GPR68 mediated IL-8 production in human pancreatic β cells
- PMID: 27166427
- PMCID: PMC4863151
- DOI: 10.1038/srep25765
Extracellular acidification stimulates GPR68 mediated IL-8 production in human pancreatic β cells
Abstract
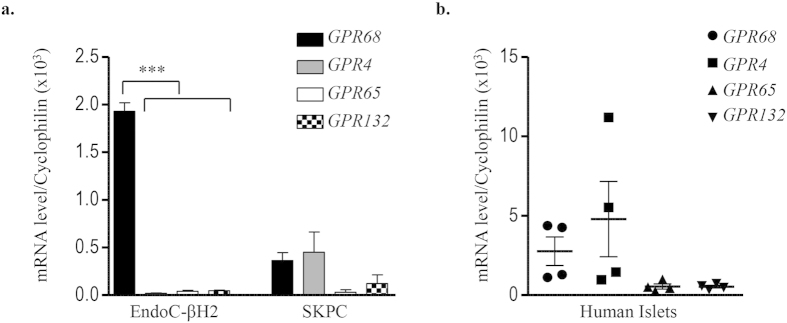
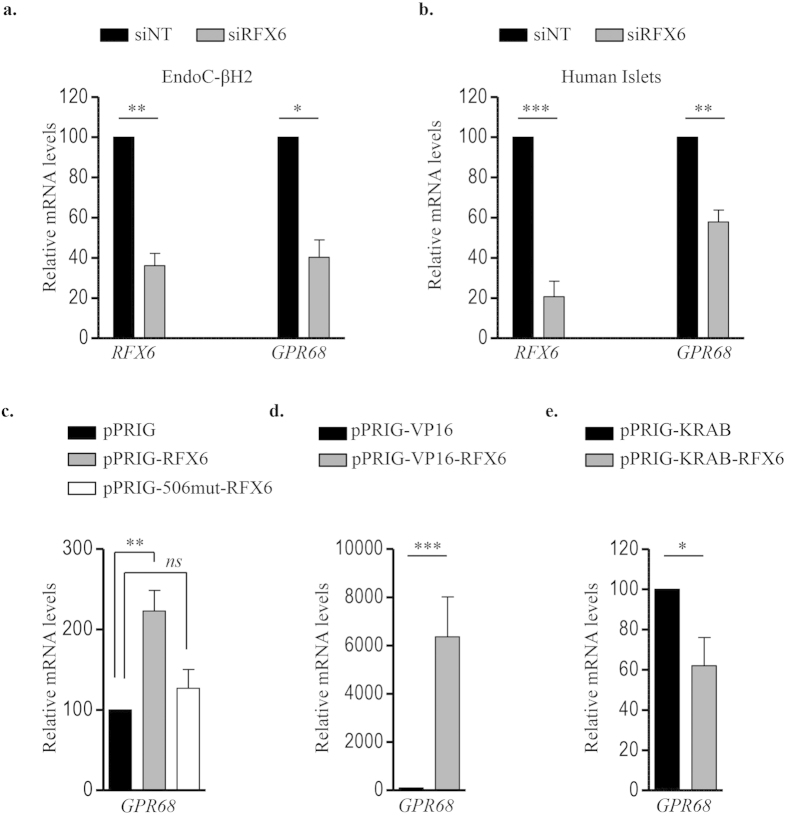
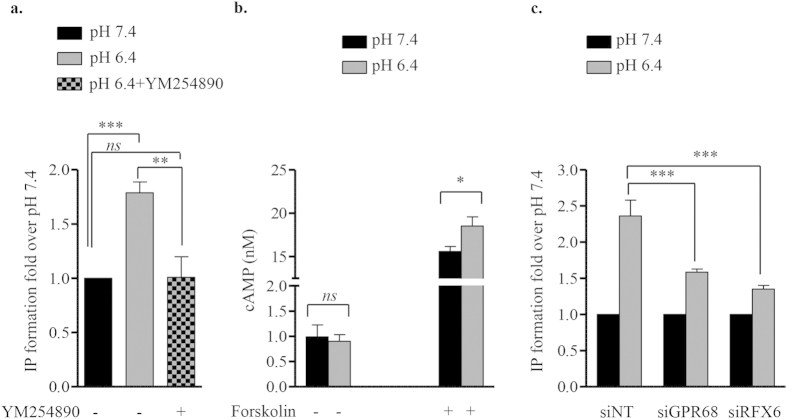
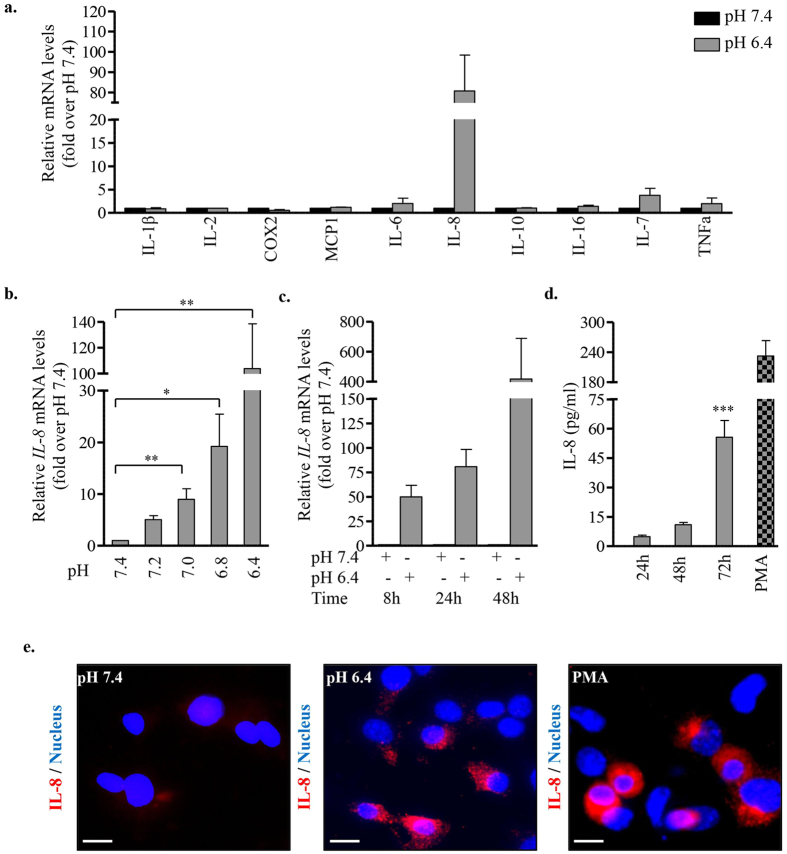
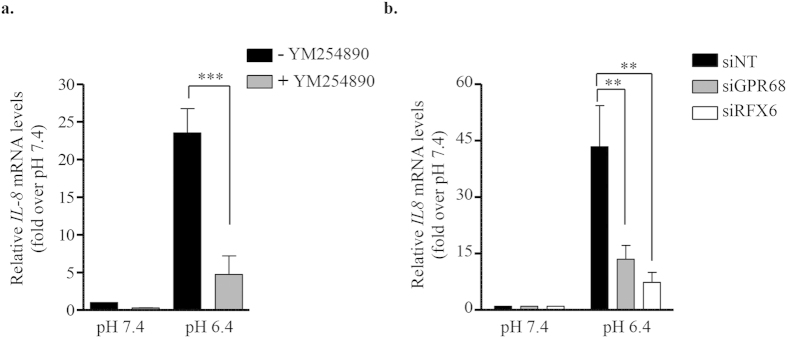
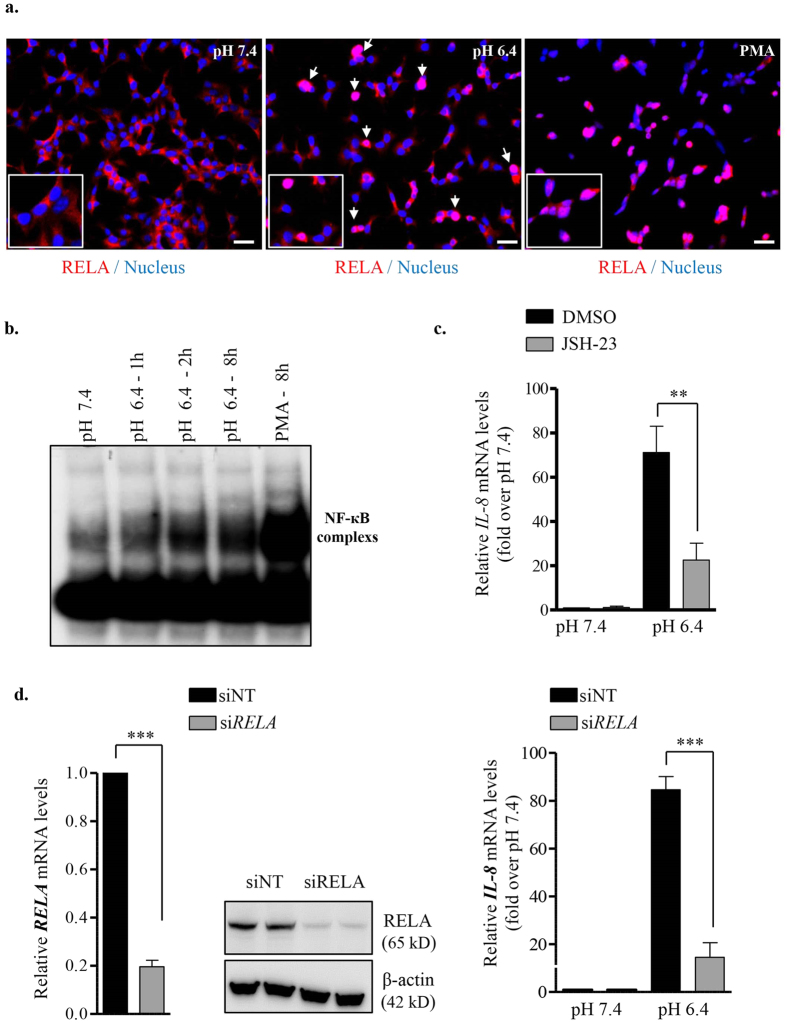
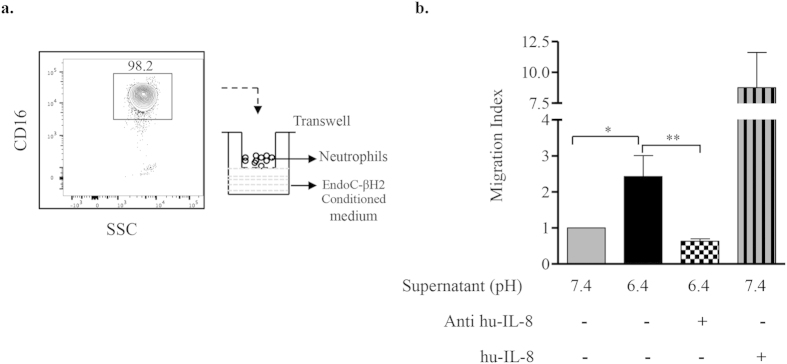
Acute or chronic metabolic complications such as diabetic ketoacidosis are often associated with extracellular acidification and pancreatic β-cell dysfunction. However, the mechanisms by which human β-cells sense and respond to acidic pH remain elusive. In this study, using the recently developed human β-cell line EndoC-βH2, we demonstrate that β-cells respond to extracellular acidification through GPR68, which is the predominant proton sensing receptor of human β-cells. Using gain- and loss-of-function studies, we provide evidence that the β-cell enriched transcription factor RFX6 is a major regulator of GPR68. Further, we show that acidic pH stimulates the production and secretion of the chemokine IL-8 by β-cells through NF-кB activation. Blocking of GPR68 or NF-кB activity severely attenuated acidification induced IL-8 production. Thus, we provide mechanistic insights into GPR68 mediated β-cell response to acidic microenvironment, which could be a new target to protect β-cell against acidosis induced inflammation.
Figures
Similar articles
-
Proton receptor GPR68 expression in dendritic-cell-like S100β-positive cells of rat anterior pituitary gland: GPR68 induces interleukin-6 gene expression in extracellular acidification.Cell Tissue Res. 2014 Nov;358(2):515-25. doi: 10.1007/s00441-014-1958-x. Epub 2014 Aug 17. Cell Tissue Res. 2014. PMID: 25129106
-
Ovarian cancer G-protein-coupled receptor 1 induces the expression of the pain mediator prostaglandin E2 in response to an acidic extracellular environment in human osteoblast-like cells.Int J Biochem Cell Biol. 2012 Nov;44(11):1937-41. doi: 10.1016/j.biocel.2012.07.015. Epub 2012 Jul 23. Int J Biochem Cell Biol. 2012. PMID: 22835475
-
Involvement of proton-sensing TDAG8 in extracellular acidification-induced inhibition of proinflammatory cytokine production in peritoneal macrophages.J Immunol. 2009 Mar 1;182(5):3243-51. doi: 10.4049/jimmunol.0803466. J Immunol. 2009. PMID: 19234222
-
Regulation of inflammation by extracellular acidification and proton-sensing GPCRs.Cell Signal. 2013 Nov;25(11):2263-71. doi: 10.1016/j.cellsig.2013.07.022. Epub 2013 Jul 31. Cell Signal. 2013. PMID: 23917207 Review.
-
Role of extracellular proton-sensing OGR1 in regulation of insulin secretion and pancreatic β-cell functions.Endocr J. 2014;61(2):101-10. doi: 10.1507/endocrj.ej13-0380. Epub 2013 Oct 1. Endocr J. 2014. PMID: 24088601 Review.
Cited by
-
Endoplasmic Reticulum-Mitochondria Crosstalk and Beta-Cell Destruction in Type 1 Diabetes.Front Immunol. 2021 Apr 16;12:669492. doi: 10.3389/fimmu.2021.669492. eCollection 2021. Front Immunol. 2021. PMID: 33936111 Free PMC article. Review.
-
Orphan GPR61, GPR62 and GPR135 receptors and the melatonin MT2 receptor reciprocally modulate their signaling functions.Sci Rep. 2017 Aug 21;7(1):8990. doi: 10.1038/s41598-017-08996-7. Sci Rep. 2017. PMID: 28827538 Free PMC article.
-
Extracellular acidification-induced CXCL8 production through a proton-sensing receptor OGR1 in human airway smooth muscle cells: a response inhibited by dexamethasone.J Inflamm (Lond). 2019 Feb 19;16:4. doi: 10.1186/s12950-019-0207-1. eCollection 2019. J Inflamm (Lond). 2019. PMID: 30828266 Free PMC article.
-
Comprehensive Assessment of GPR68 Expression in Normal and Neoplastic Human Tissues Using a Novel Rabbit Monoclonal Antibody.Int J Mol Sci. 2019 Oct 23;20(21):5261. doi: 10.3390/ijms20215261. Int J Mol Sci. 2019. PMID: 31652823 Free PMC article.
-
Mitchell-Riley Syndrome: Improving Clinical Outcomes and Searching for Functional Impact of RFX-6 Mutations.Front Endocrinol (Lausanne). 2022 Jun 22;13:802351. doi: 10.3389/fendo.2022.802351. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 35813646 Free PMC article.
References
-
- Griffith L. G. & Swartz M. a. Capturing complex 3 D tissue physiology in vitro. Nat. Rev. Mol. Cell Biol. 7, 211–24 (2006). - PubMed
-
- Lardner A. The effects of extracellular pH on immune function. J. Leukoc. Biol. 69, 522–530 (2001). - PubMed
-
- Wemmie J. a., Price M. P. & Welsh M. J. Acid-sensing ion channels: advances, questions and therapeutic opportunities. Trends Neurosci. 29, 578–586 (2006). - PubMed
-
- Okajima F. Regulation of inflammation by extracellular acidification and proton-sensing {GPCRs.}. Cell. Signal. 25, 2263–2271 (2013). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
