

Protonation Dynamics on Lipid Nanodiscs: Influence of the Membrane Surface Area and External Buffers
- PMID: 27166807
- PMCID: PMC4939474
- DOI: 10.1016/j.bpj.2016.03.035
Protonation Dynamics on Lipid Nanodiscs: Influence of the Membrane Surface Area and External Buffers
Abstract
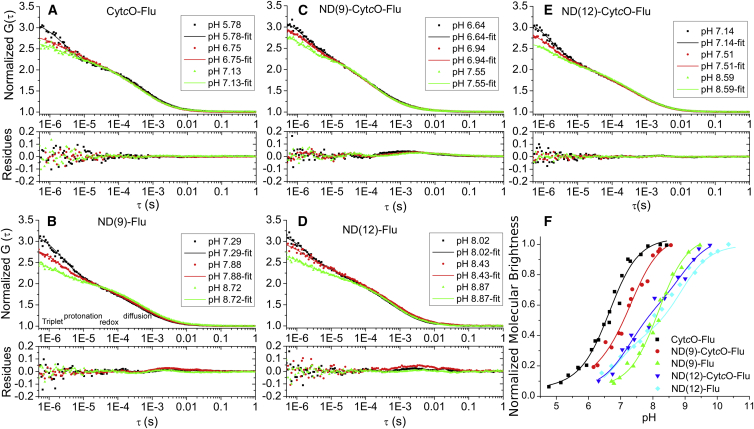
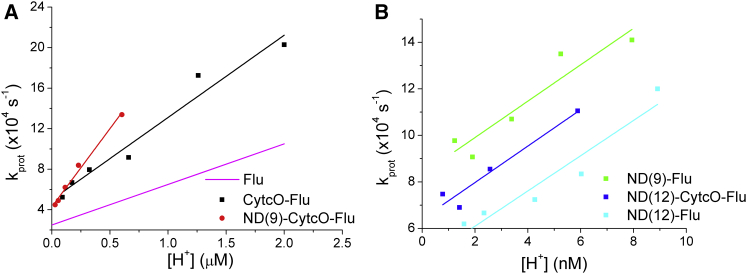
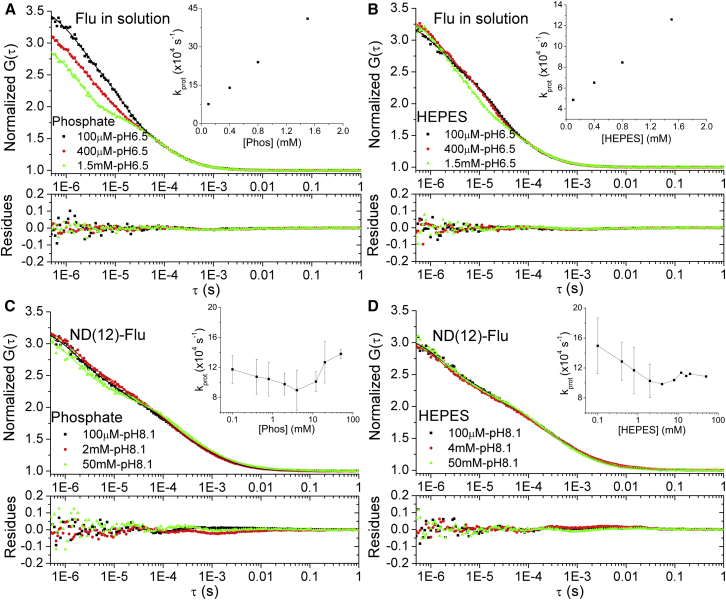
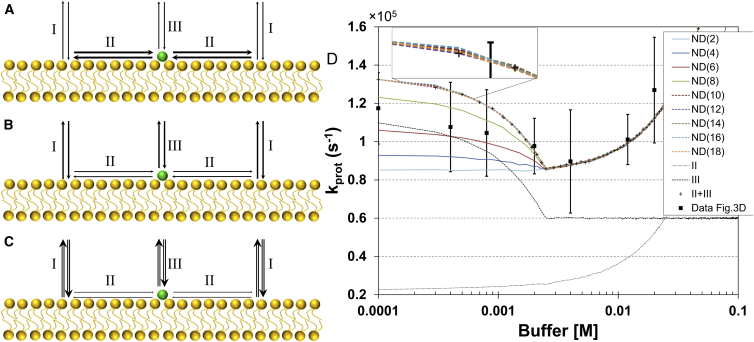
Lipid membrane surfaces can act as proton-collecting antennae, accelerating proton uptake by membrane-bound proton transporters. We investigated this phenomenon in lipid nanodiscs (NDs) at equilibrium on a local scale, analyzing fluorescence fluctuations of individual pH-sensitive fluorophores at the membrane surface by fluorescence correlation spectroscopy (FCS). The protonation rate of the fluorophores was ∼100-fold higher when located at 9- and 12-nm diameter NDs, compared to when in solution, indicating that the proton-collecting antenna effect is maximal already for a membrane area of ∼60 nm(2). Fluorophore-labeled cytochrome c oxidase displayed a similar increase when reconstituted in 12 nm NDs, but not in 9 nm NDs, i.e., an acceleration of the protonation rate at the surface of cytochrome c oxidase is found when the lipid area surrounding the protein is larger than 80 nm(2), but not when below 30 nm(2). We also investigated the effect of external buffers on the fluorophore proton exchange rates at the ND membrane-water interfaces. With increasing buffer concentrations, the proton exchange rates were found to first decrease and then, at millimolar buffer concentrations, to increase. Monte Carlo simulations, based on a simple kinetic model of the proton exchange at the membrane-water interface, and using rate parameter values determined in our FCS experiments, could reconstruct both the observed membrane-size and the external buffer dependence. The FCS data in combination with the simulations indicate that the local proton diffusion coefficient along a membrane is ∼100 times slower than that observed over submillimeter distances by proton-pulse experiments (Ds ∼ 10(-5)cm(2)/s), and support recent theoretical studies showing that proton diffusion along membrane surfaces is time- and length-scale dependent.
Copyright © 2016 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
Proton Dynamics at the Membrane Surface.Biophys J. 2016 May 10;110(9):1909-11. doi: 10.1016/j.bpj.2016.04.001. Biophys J. 2016. PMID: 27166799 Free PMC article. No abstract available.
References
-
- Sackes V., Marantz Y., Gutman M. The dynamic feature of the proton collecting antenna of a protein surface. Biochim. Biophys. Acta. 1998;1365:232–240.
-
- Ädelroth P., Brzezinski P. Surface-mediated proton-transfer reactions in membrane-bound proteins. Biochim. Biophys. Acta. 2004;1655:102–115. - PubMed
-
- Heberle J. Proton transfer reactions across bacteriorhodopsin and along the membrane. Biochim. Biophys. Acta. 2000;1458:135–147. - PubMed
-
- Mulkidjanian A.Y., Heberle J., Cherepanov D.A. Protons @ interfaces: implications for biological energy conversion. Biochim. Biophys. Acta. 2006;1757:913–930. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
