

Progesterone-Dependent Induction of Phospholipase C-Related Catalytically Inactive Protein 1 (PRIP-1) in Decidualizing Human Endometrial Stromal Cells
- PMID: 27167772
- PMCID: PMC4972893
- DOI: 10.1210/en.2015-1914
Progesterone-Dependent Induction of Phospholipase C-Related Catalytically Inactive Protein 1 (PRIP-1) in Decidualizing Human Endometrial Stromal Cells
Abstract
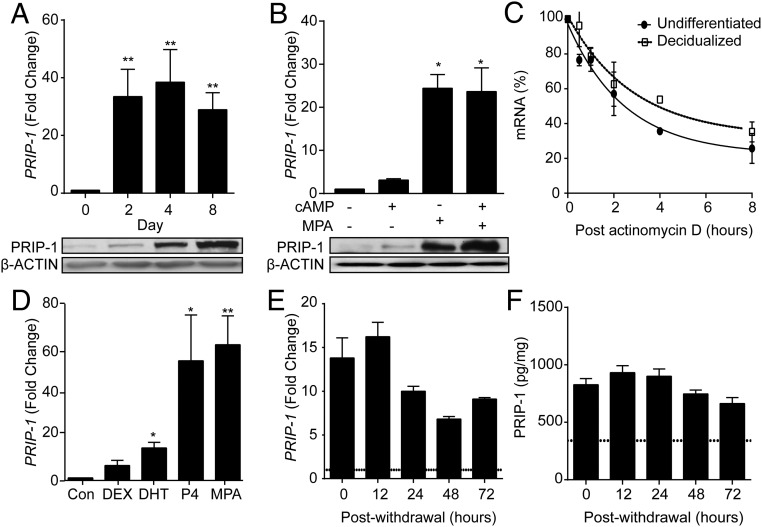
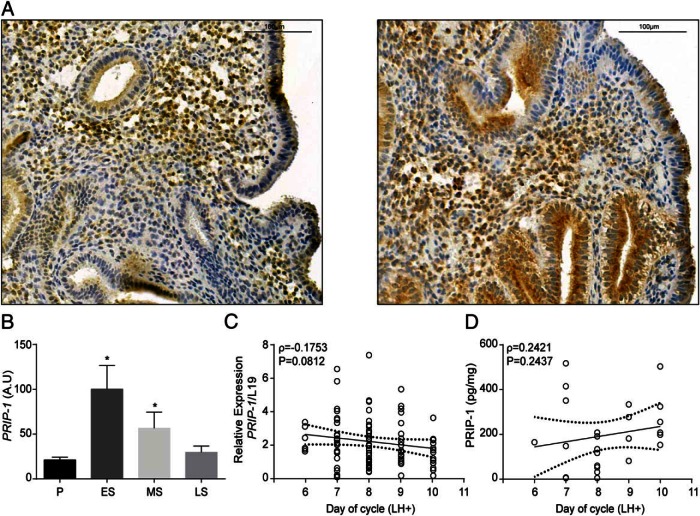
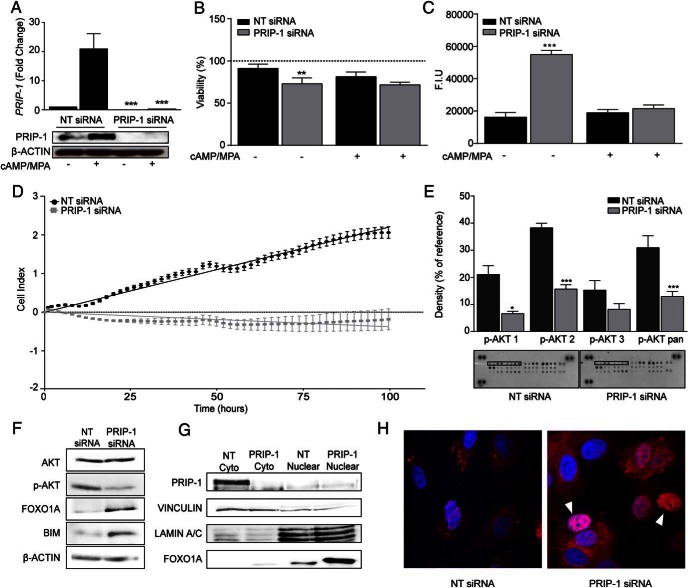
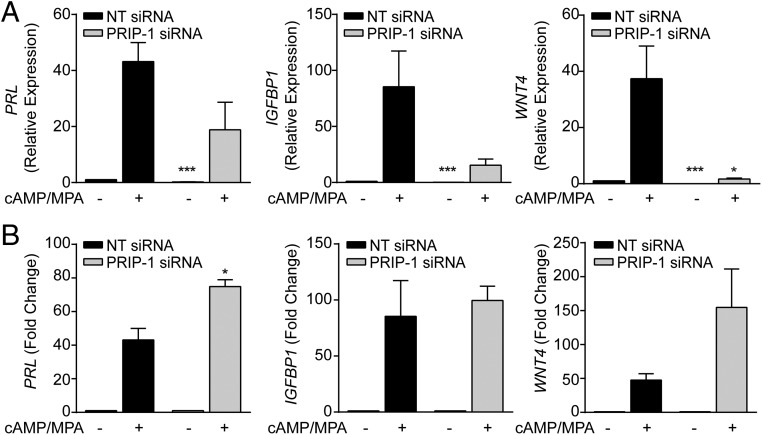
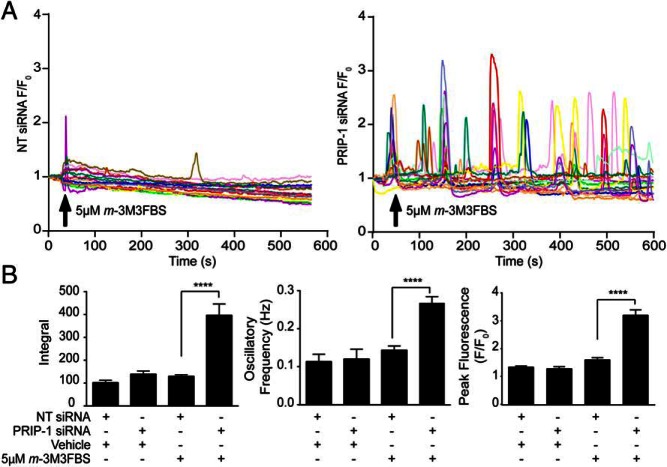
Decidualization denotes the transformation of endometrial stromal cells into specialized decidual cells. In pregnancy, decidual cells form a protective matrix around the implanting embryo, enabling coordinated trophoblast invasion and formation of a functional placenta. Continuous progesterone (P4) signaling renders decidual cells resistant to various environmental stressors, whereas withdrawal inevitably triggers tissue breakdown and menstruation or miscarriage. Here, we show that PLCL1, coding phospholipase C (PLC)-related catalytically inactive protein 1 (PRIP-1), is highly induced in response to P4 signaling in decidualizing human endometrial stromal cells (HESCs). Knockdown experiments in undifferentiated HESCs revealed that PRIP-1 maintains basal phosphoinositide 3-kinase/Protein kinase B activity, which in turn prevents illicit nuclear translocation of the transcription factor forkhead box protein O1 and induction of the apoptotic activator BIM. By contrast, loss of this scaffold protein did not compromise survival of decidual cells. PRIP-1 knockdown did also not interfere with the responsiveness of HESCs to deciduogenic cues, although the overall expression of differentiation markers, such as PRL, IGFBP1, and WNT4, was blunted. Finally, we show that PRIP-1 in decidual cells uncouples PLC activation from intracellular Ca(2+) release by attenuating inositol 1,4,5-trisphosphate signaling. In summary, PRIP-1 is a multifaceted P4-inducible scaffold protein that gates the activity of major signal transduction pathways in the endometrium. It prevents apoptosis of proliferating stromal cells and contributes to the relative autonomy of decidual cells by silencing PLC signaling downstream of Gq protein-coupled receptors.
Figures
References
-
- Gellersen B, Brosens JJ. Cyclic decidualization of the human endometrium in reproductive health and failure. Endocr Rev. 2014;35:851–905. - PubMed
-
- Bernardini L, Moretti-Rojas I, Brush M, Rojas FJ, Balmaceda JP. Changes in expression of adenyl cyclase activity in human endometrium during hormone replacement therapy and ovarian stimulation. Mol Hum Reprod. 1999;5:955–960. - PubMed
-
- Brosens JJ, Hayashi N, White JO. Progesterone receptor regulates decidual prolactin expression in differentiating human endometrial stromal cells. Endocrinology. 1999;140:4809–4820. - PubMed
-
- Evans J, Salamonsen LA. Inflammation, leukocytes and menstruation. Rev Endocr Metab Disord. 2012;13:277–288. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
