

Virus Infections Incite Pain Hypersensitivity by Inducing Indoleamine 2,3 Dioxygenase
- PMID: 27168185
- PMCID: PMC4863962
- DOI: 10.1371/journal.ppat.1005615
Virus Infections Incite Pain Hypersensitivity by Inducing Indoleamine 2,3 Dioxygenase
Abstract
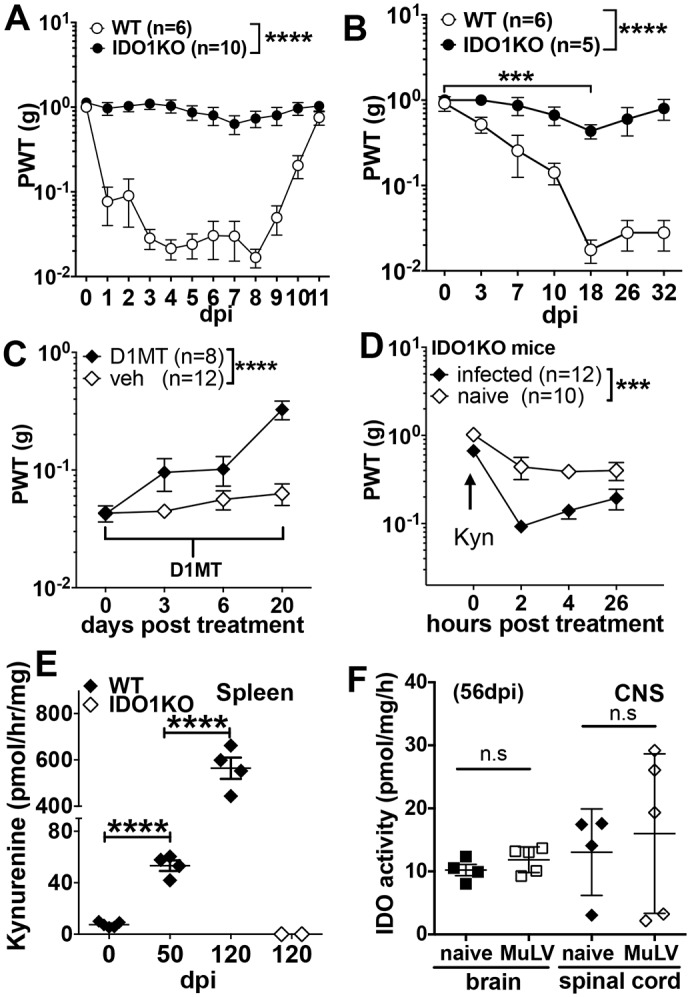
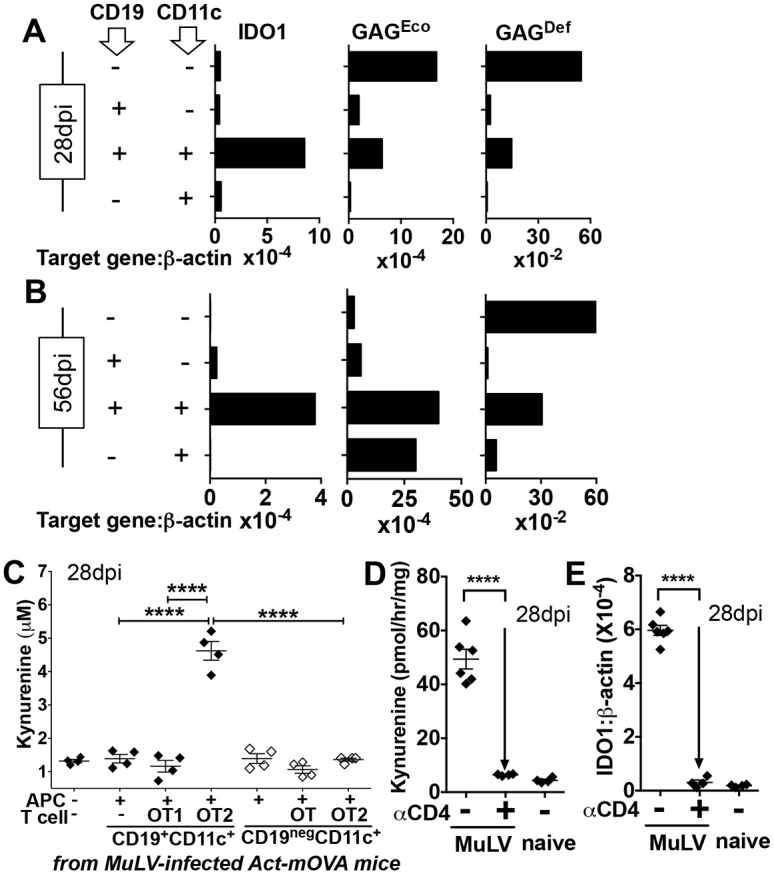
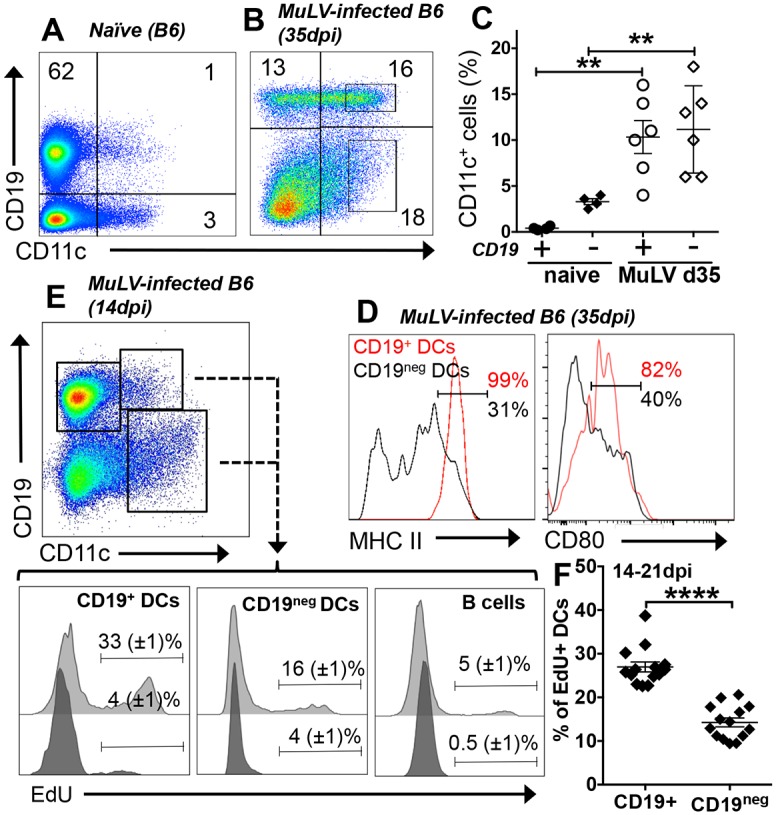
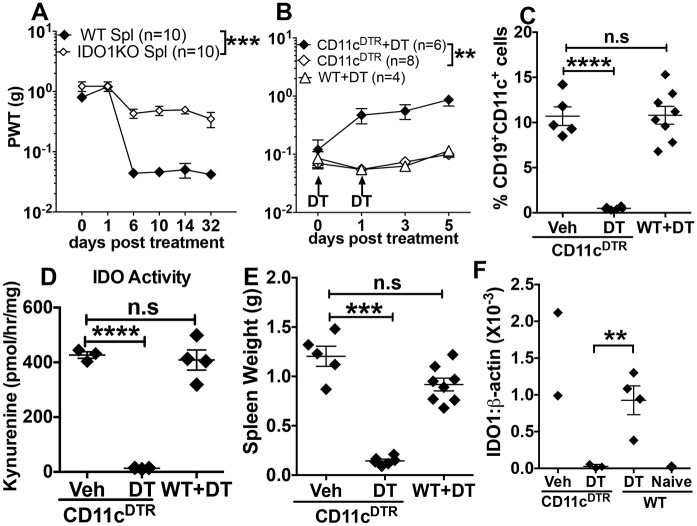
Increased pain sensitivity is a comorbidity associated with many clinical diseases, though the underlying causes are poorly understood. Recently, chronic pain hypersensitivity in rodents treated to induce chronic inflammation in peripheral tissues was linked to enhanced tryptophan catabolism in brain mediated by indoleamine 2,3 dioxygenase (IDO). Here we show that acute influenza A virus (IAV) and chronic murine leukemia retrovirus (MuLV) infections, which stimulate robust IDO expression in lungs and lymphoid tissues, induced acute or chronic pain hypersensitivity, respectively. In contrast, virus-induced pain hypersensitivity did not manifest in mice lacking intact IDO1 genes. Spleen IDO activity increased markedly as MuLV infections progressed, while IDO1 expression was not elevated significantly in brain or spinal cord (CNS) tissues. Moreover, kynurenine (Kyn), a tryptophan catabolite made by cells expressing IDO, incited pain hypersensitivity in uninfected IDO1-deficient mice and Kyn potentiated pain hypersensitivity due to MuLV infection. MuLV infection stimulated selective IDO expression by a discreet population of spleen cells expressing both B cell (CD19) and dendritic cell (CD11c) markers (CD19+ DCs). CD19+ DCs were more susceptible to MuLV infection than B cells or conventional (CD19neg) DCs, proliferated faster than B cells from early stages of MuLV infection and exhibited mature antigen presenting cell (APC) phenotypes, unlike conventional (CD19neg) DCs. Moreover, interactions with CD4 T cells were necessary to sustain functional IDO expression by CD19+ DCs in vitro and in vivo. Splenocytes from MuLV-infected IDO1-sufficient mice induced pain hypersensitivity in uninfected IDO1-deficient recipient mice, while selective in vivo depletion of DCs alleviated pain hypersensitivity in MuLV-infected IDO1-sufficient mice and led to rapid reduction in splenomegaly, a hallmark of MuLV immune pathogenesis. These findings reveal critical roles for CD19+ DCs expressing IDO in host responses to MuLV infection that enhance pain hypersensitivity and cause immune pathology. Collectively, our findings support the hypothesis elevated IDO activity in non-CNS due to virus infections causes pain hypersensitivity mediated by Kyn. Previously unappreciated links between host immune responses to virus infections and pain sensitivity suggest that IDO inhibitors may alleviate heightened pain sensitivity during infections.
Conflict of interest statement
I have read the journal's policy and the authors of this manuscript have the following competing interests: ALM and DHM serve as consultants for NewLink Genetics Inc. and receive remuneration from this source. The authors declare no additional competing financial interests. This does not alter our adherence to all PLOS policies on sharing data and materials.
Figures
References
-
- Sindic CJ. Infectious neuropathies. Current opinion in neurology. 2013;26(5):510–5. Epub 2013/08/16. . - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
