

Limitations of Neural Map Topography for Decoding Spatial Information
- PMID: 27170134
- PMCID: PMC6601797
- DOI: 10.1523/JNEUROSCI.0385-16.2016
Limitations of Neural Map Topography for Decoding Spatial Information
Abstract
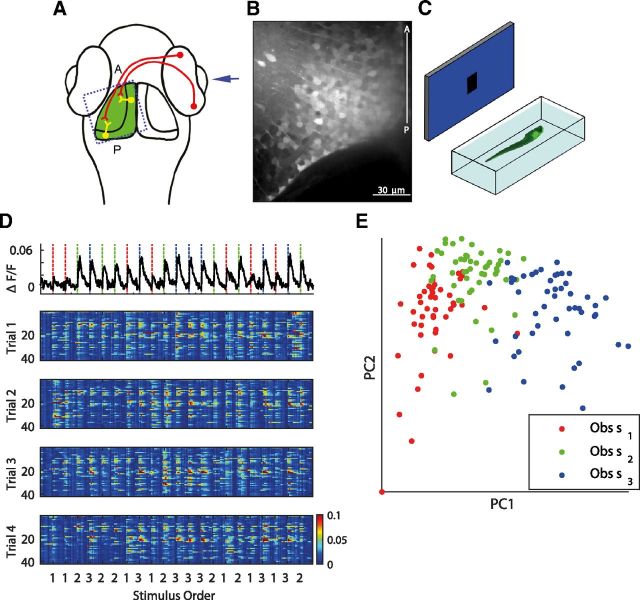
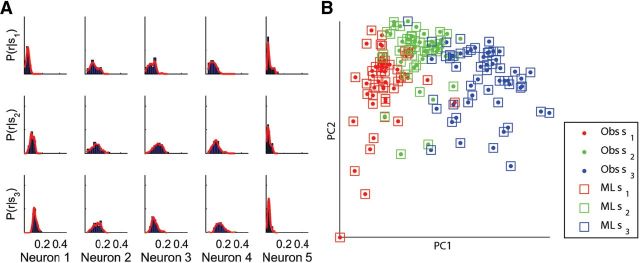
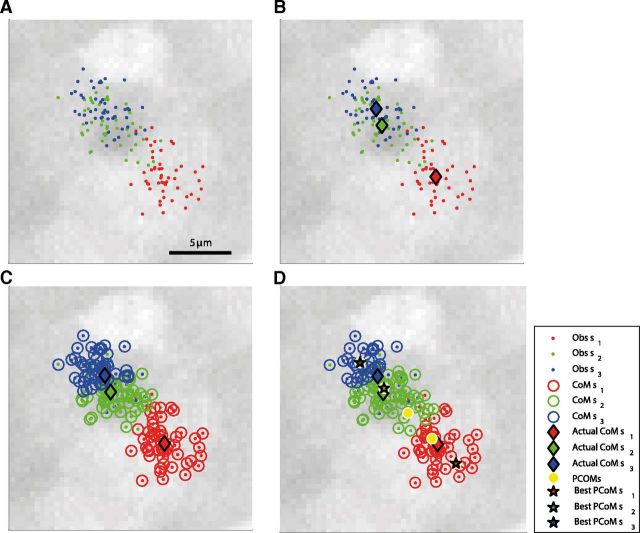
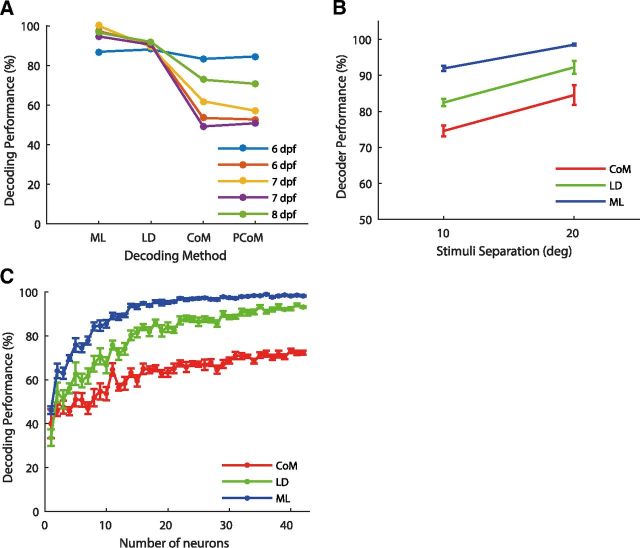
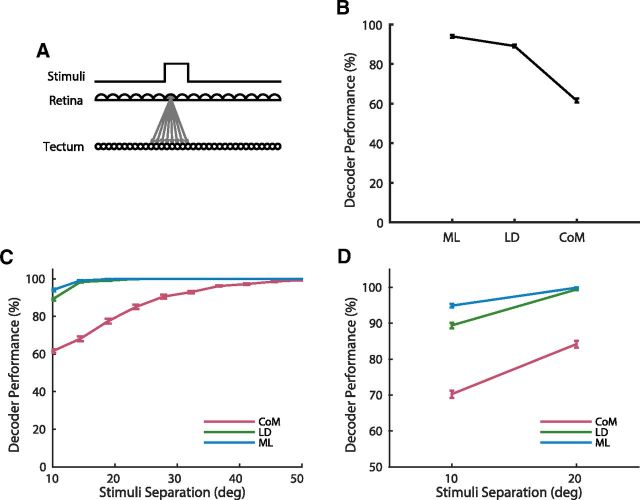
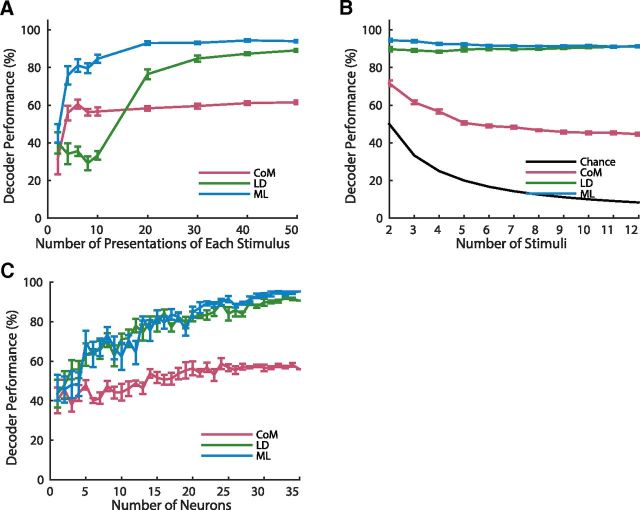
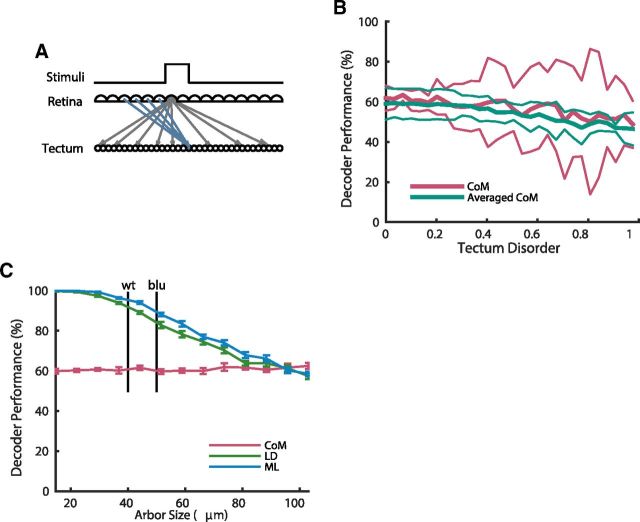
Topographic maps are common throughout the nervous system, yet their functional role is still unclear. In particular, whether they are necessary for decoding sensory stimuli is unknown. Here we examined this question by recording population activity at the cellular level from the larval zebrafish tectum in response to visual stimuli at three closely spaced locations in the visual field. Due to map imprecision, nearby stimulus locations produced intermingled tectal responses, and decoding based on map topography yielded an accuracy of only 64%. In contrast, maximum likelihood decoding of stimulus location based on the statistics of the evoked activity, while ignoring any information about the locations of neurons in the map, yielded an accuracy close to 100%. A simple computational model of the zebrafish visual system reproduced these results. Although topography is a useful initial decoding strategy, we suggest it may be replaced by better methods following visual experience.
Significance statement: A very common feature of brain wiring is that neighboring points on a sensory surface (eg, the retina) are connected to neighboring points in the brain. It is often assumed that this "topography" of wiring is essential for decoding sensory stimuli. However, here we show in the developing zebrafish that topographic decoding performs very poorly compared with methods that do not rely on topography. This suggests that, although wiring topography could provide a starting point for decoding at a very early stage in development, it may be replaced by more accurate methods as the animal gains experience of the world.
Keywords: computational model; sensory decoding; topographic map; zebrafish.
Copyright © 2016 the authors 0270-6474/16/365385-12$15.00/0.
Figures
References
-
- Beyer JE. Feeding success of clupeoid fish larvae and stochastic thinking. Dana. 1980;1:65–91.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases