

Virus-derived small RNAs: molecular footprints of host-pathogen interactions
- PMID: 27170499
- PMCID: PMC7169819
- DOI: 10.1002/wrna.1361
Virus-derived small RNAs: molecular footprints of host-pathogen interactions
Abstract
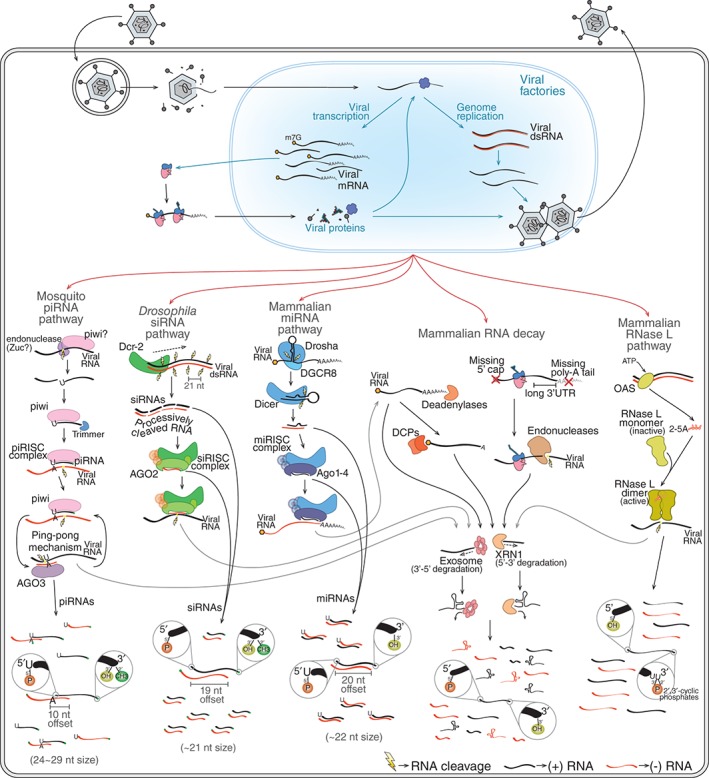
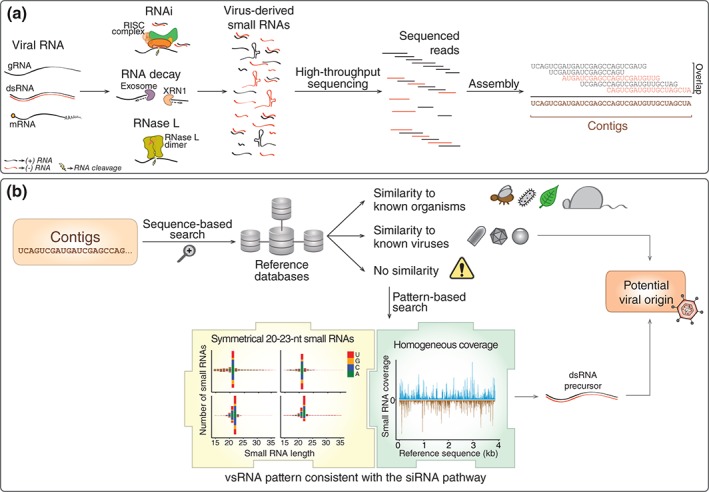
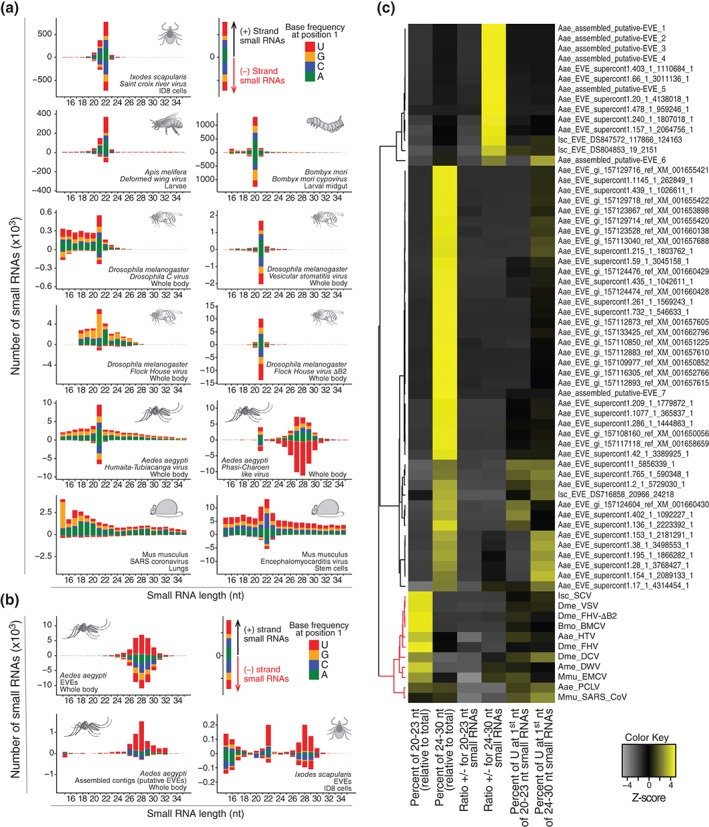
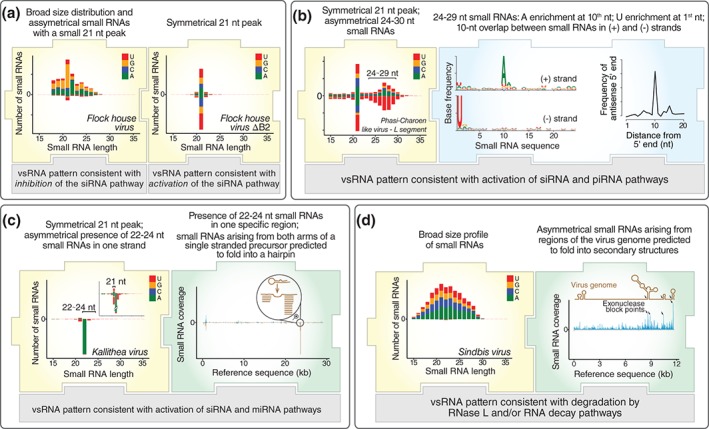
Viruses are obligatory intracellular parasites that require the host machinery to replicate. During their replication cycle, viral RNA intermediates can be recognized and degraded by different antiviral mechanisms that include RNA decay, RNA interference, and RNase L pathways. As a consequence of viral RNA degradation, infected cells can accumulate virus-derived small RNAs at high levels compared to cellular molecules. These small RNAs are imprinted with molecular characteristics that reflect their origin. First, small RNAs can be used to reconstruct viral sequences and identify the virus from which they originated. Second, other molecular features of small RNAs such as size, polarity, and base preferences depend on the type of viral substrate and host mechanism of degradation. Thus, the pattern of small RNAs generated in infected cells can be used as a molecular footprint to identify and characterize viruses independent on sequence homology searches against known references. Hence, sequencing of small RNAs obtained from infected cells enables virus discovery and characterization using both sequence-dependent strategies and novel pattern-based approaches. Recent studies are helping unlock the full application of small RNA sequencing for virus discovery and characterization. WIREs RNA 2016, 7:824-837. doi: 10.1002/wrna.1361 For further resources related to this article, please visit the WIREs website.
© 2016 Wiley Periodicals, Inc.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
