

Pollen Killer Gene S35 Function Requires Interaction with an Activator That Maps Close to S24, Another Pollen Killer Gene in Rice
- PMID: 27172610
- PMCID: PMC4856096
- DOI: 10.1534/g3.116.027573
Pollen Killer Gene S35 Function Requires Interaction with an Activator That Maps Close to S24, Another Pollen Killer Gene in Rice
Abstract
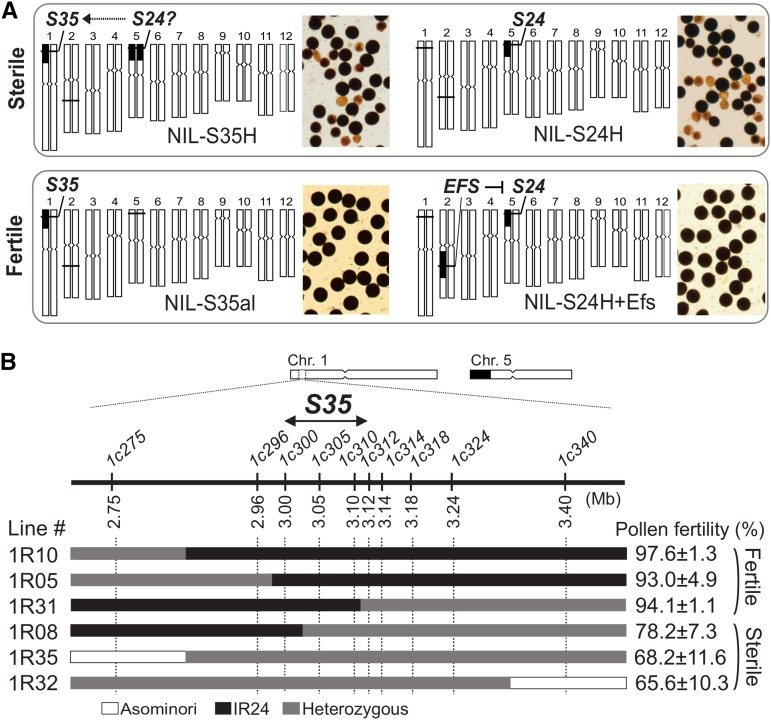
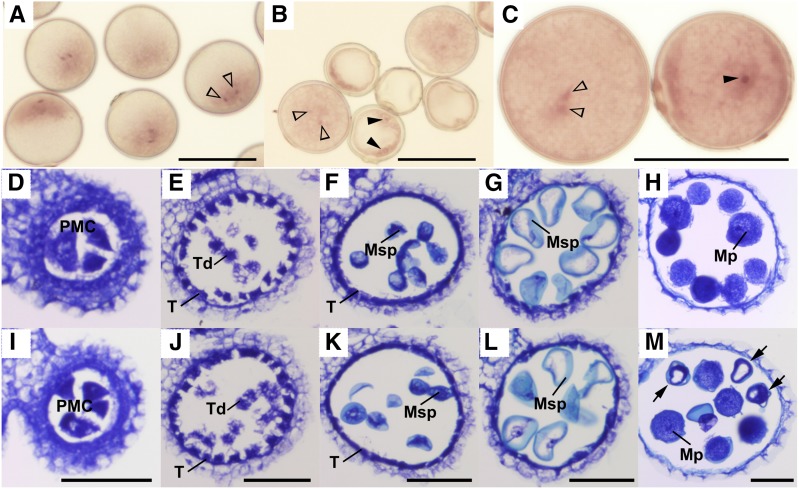
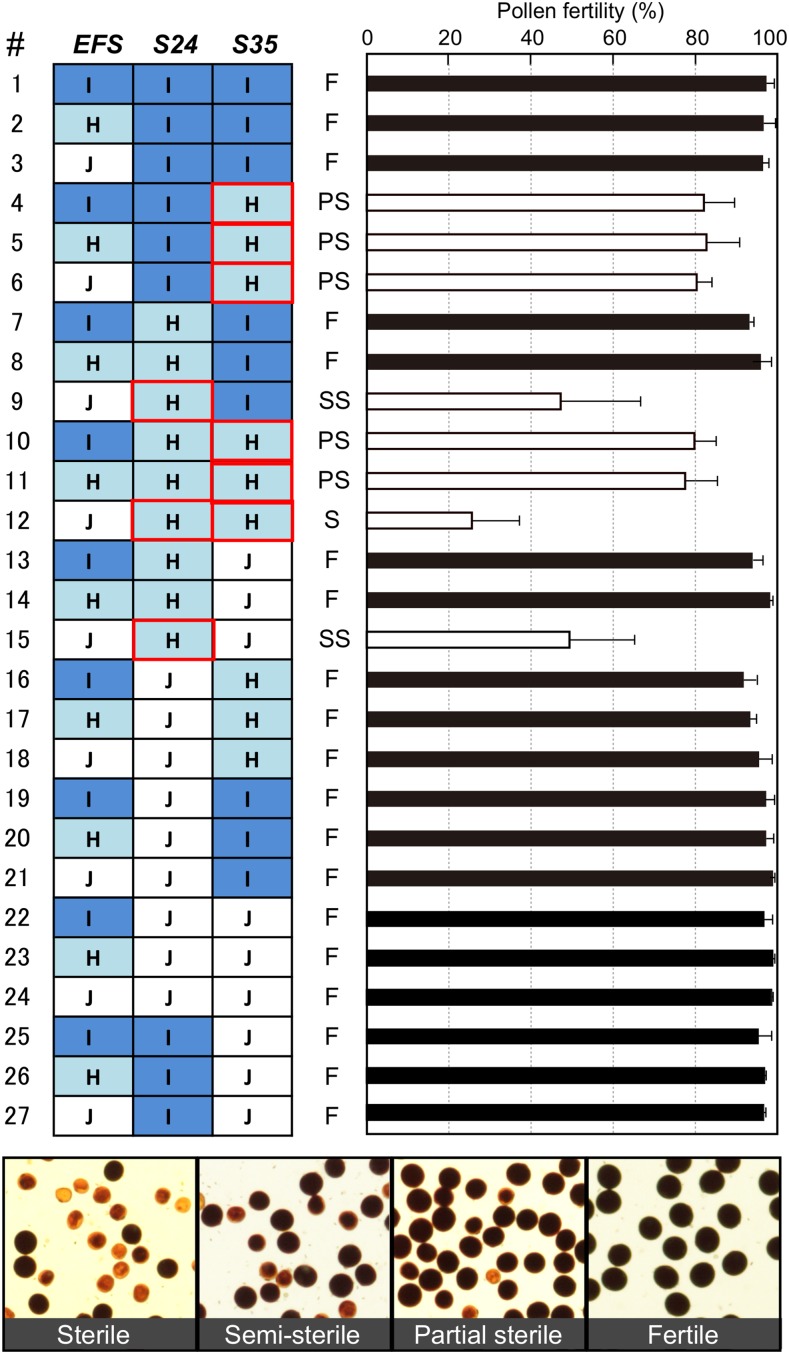
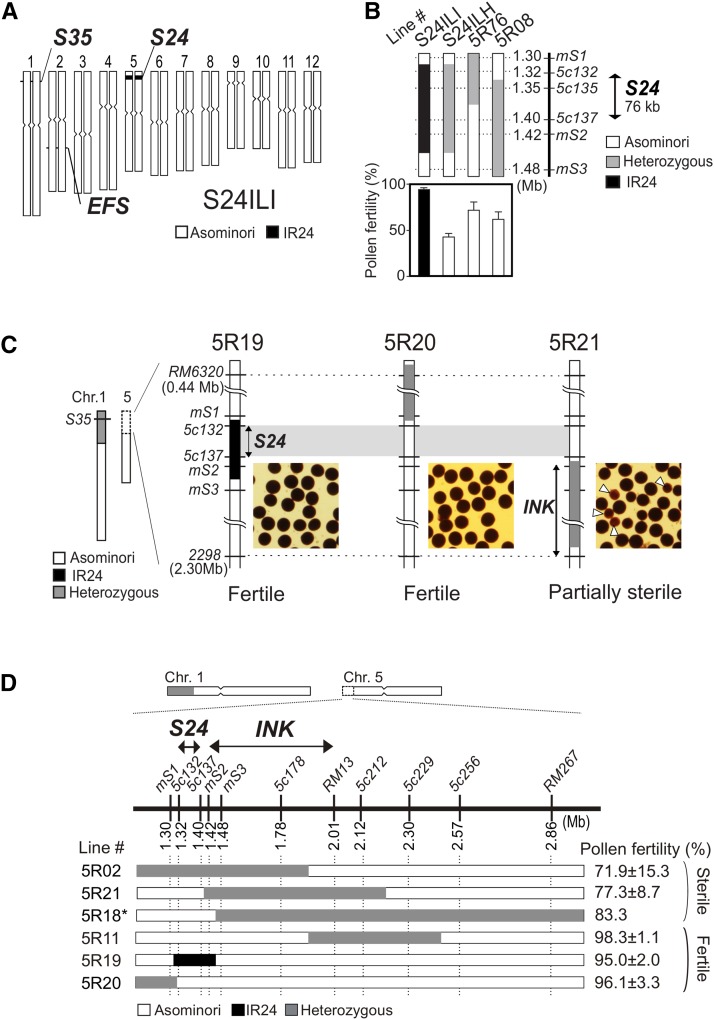
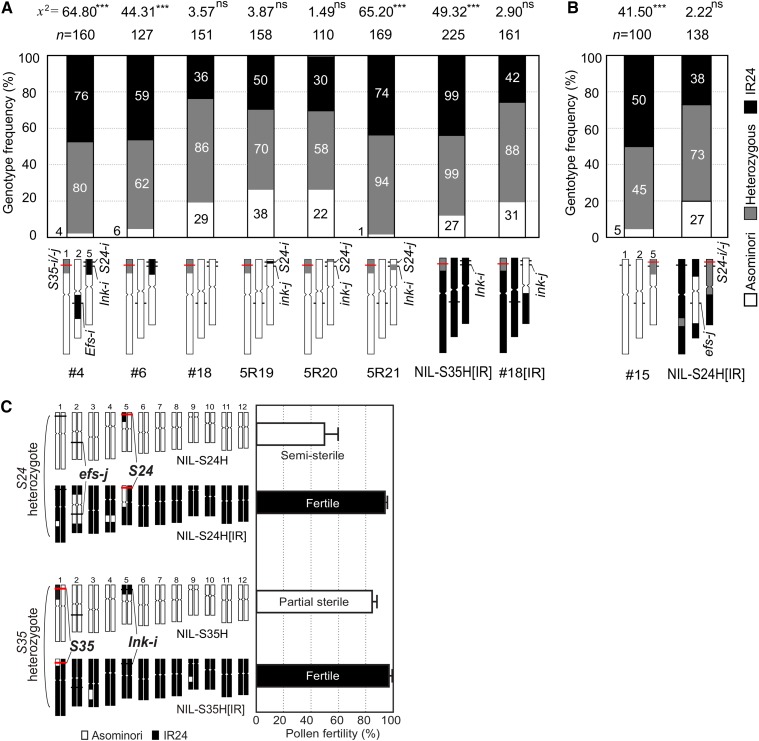
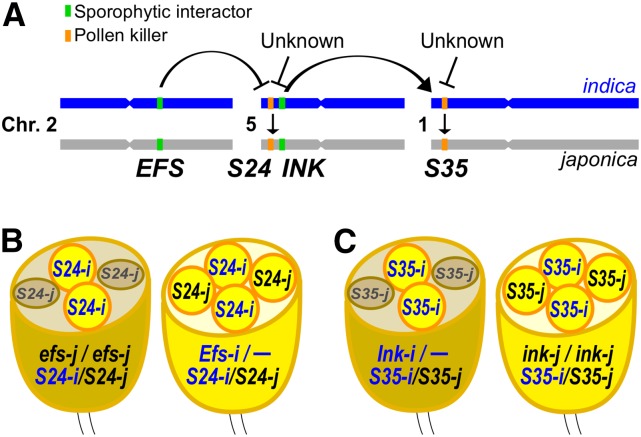
Pollen killer genes disable noncarrier pollens, and are responsible for male sterility and segregation distortion in hybrid populations of distantly related plant species. The genetic networks and the molecular mechanisms underlying the pollen killer system remain largely unknown. Two pollen killer genes, S24 and S35, have been found in an intersubspecific cross of Oryza sativa ssp. indica and japonica The effect of S24 is counteracted by an unlinked locus EFS Additionally, S35 has been proposed to interact with S24 to induce pollen sterility. These genetic interactions are suggestive of a single S24-centric genetic pathway (EFS-S24-S35) for the pollen killer system. To examine this hypothetical genetic pathway, the S35 and the S24 regions were further characterized and genetically dissected in this study. Our results indicated that S35 causes pollen sterility independently of both the EFS and S24 genes, but is dependent on a novel gene close to the S24 locus, named incentive for killing pollen (INK). We confirmed the phenotypic effect of the INK gene separately from the S24 gene, and identified the INK locus within an interval of less than 0.6 Mb on rice chromosome 5. This study characterized the genetic effect of the two independent genetic pathways of INK-S35 and EFS-S24 in indica-japonica hybrid progeny. Our results provide clear evidence that hybrid male sterility in rice is caused by several pollen killer networks with multiple factors positively and negatively regulating pollen killer genes.
Keywords: Oryza sativa; epistasis; pollen killer; reproductive isolation.
Copyright © 2016 Kubo et al.
Figures
Similar articles
-
A novel epistatic interaction at two loci causing hybrid male sterility in an inter-subspecific cross of rice (Oryza sativa L.).Genes Genet Syst. 2008 Dec;83(6):443-53. doi: 10.1266/ggs.83.443. Genes Genet Syst. 2008. PMID: 19282622
-
Hybrid male sterility in rice is due to epistatic interactions with a pollen killer locus.Genetics. 2011 Nov;189(3):1083-92. doi: 10.1534/genetics.111.132035. Epub 2011 Aug 25. Genetics. 2011. PMID: 21868603 Free PMC article.
-
Genetic characterization and fine mapping of S25, a hybrid male sterility gene, on rice chromosome 12.Genes Genet Syst. 2018 Feb 10;92(4):205-212. doi: 10.1266/ggs.17-00012. Epub 2017 Jun 30. Genes Genet Syst. 2018. PMID: 28674278
-
Hybrid sterility in plant: stories from rice.Curr Opin Plant Biol. 2010 Apr;13(2):186-92. doi: 10.1016/j.pbi.2010.01.002. Epub 2010 Feb 10. Curr Opin Plant Biol. 2010. PMID: 20153244 Review.
-
Molecular mechanisms of hybrid sterility in rice.Sci China Life Sci. 2019 Jun;62(6):737-743. doi: 10.1007/s11427-019-9531-7. Epub 2019 May 21. Sci China Life Sci. 2019. PMID: 31119561 Review.
Cited by
-
Understanding the Nature of Hybrid Sterility and Divergence of Asian Cultivated Rice.Front Plant Sci. 2022 Jun 27;13:908342. doi: 10.3389/fpls.2022.908342. eCollection 2022. Front Plant Sci. 2022. PMID: 35832226 Free PMC article. Review.
-
Recurrent breakdown and rebalance of segregation distortion in the genomes: battle for the transmission advantage.aBIOTECH. 2020 Jul 8;1(4):246-254. doi: 10.1007/s42994-020-00023-0. eCollection 2020 Oct. aBIOTECH. 2020. PMID: 36304131 Free PMC article. Review.
-
Novel patterns of expression and recruitment of new genes on the t-haplotype, a mouse selfish chromosome.Proc Biol Sci. 2022 Feb 9;289(1968):20211985. doi: 10.1098/rspb.2021.1985. Epub 2022 Feb 9. Proc Biol Sci. 2022. PMID: 35135349 Free PMC article.
-
Influence of Gender Bias on Distribution of Hybrid Sterility in Rice.Front Plant Sci. 2022 Jul 12;13:898206. doi: 10.3389/fpls.2022.898206. eCollection 2022. Front Plant Sci. 2022. PMID: 35903237 Free PMC article. Review.
-
An overview on reproductive isolation in Oryza sativa complex.AoB Plants. 2018 Nov 22;10(6):ply060. doi: 10.1093/aobpla/ply060. eCollection 2018 Dec. AoB Plants. 2018. PMID: 30538811 Free PMC article. Review.
References
-
- Burt A., Trivers R., 2009. Genes in conflict: the biology of selfish genetic elements, Harvard University Press, Cambridge, MA.
-
- Chin J. H., Chu S.-H., Jiang W., Cho Y.-I., Basyirin R., et al. , 2011. Identification of QTLs for hybrid fertility in inter-subspecific crosses of rice (Oryza sativa L.). Genes Genomics 33: 39–48.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous