

Evidence that phytochrome functions as a protein kinase in plant light signalling
- PMID: 27173885
- PMCID: PMC4869175
- DOI: 10.1038/ncomms11545
Evidence that phytochrome functions as a protein kinase in plant light signalling
Abstract
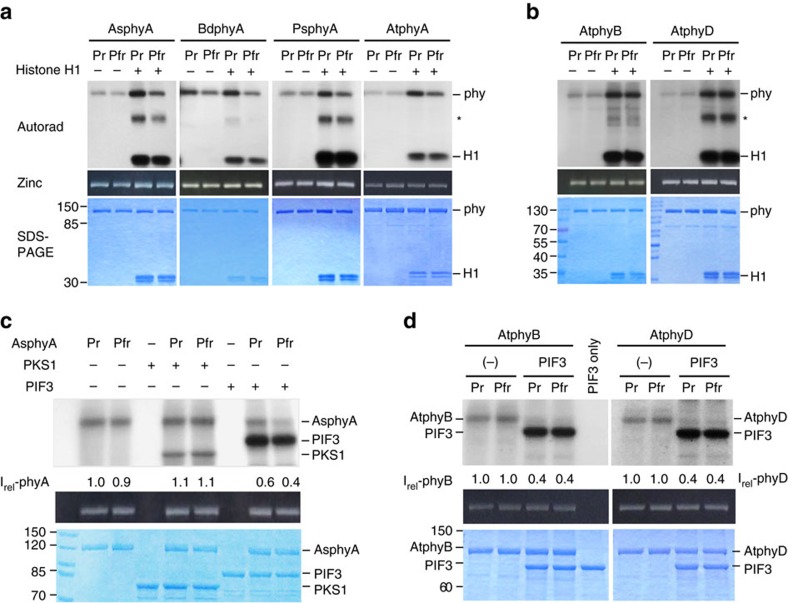
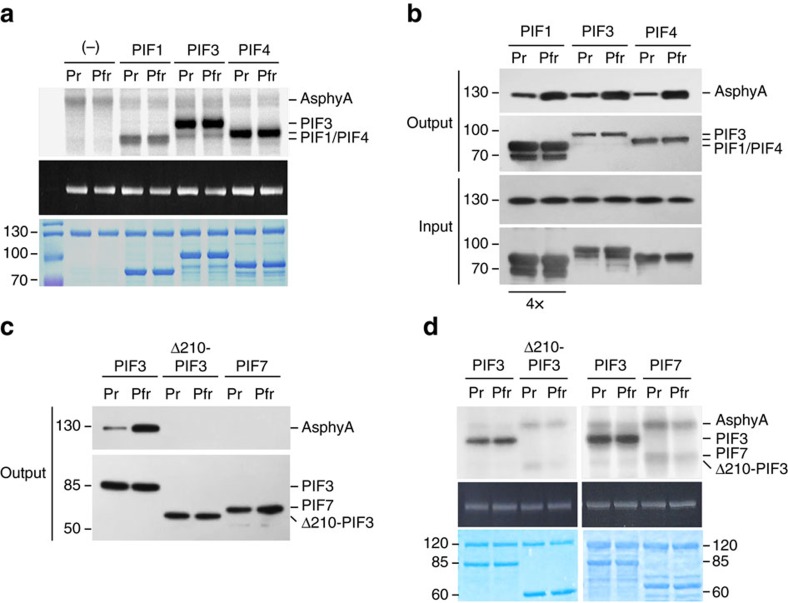
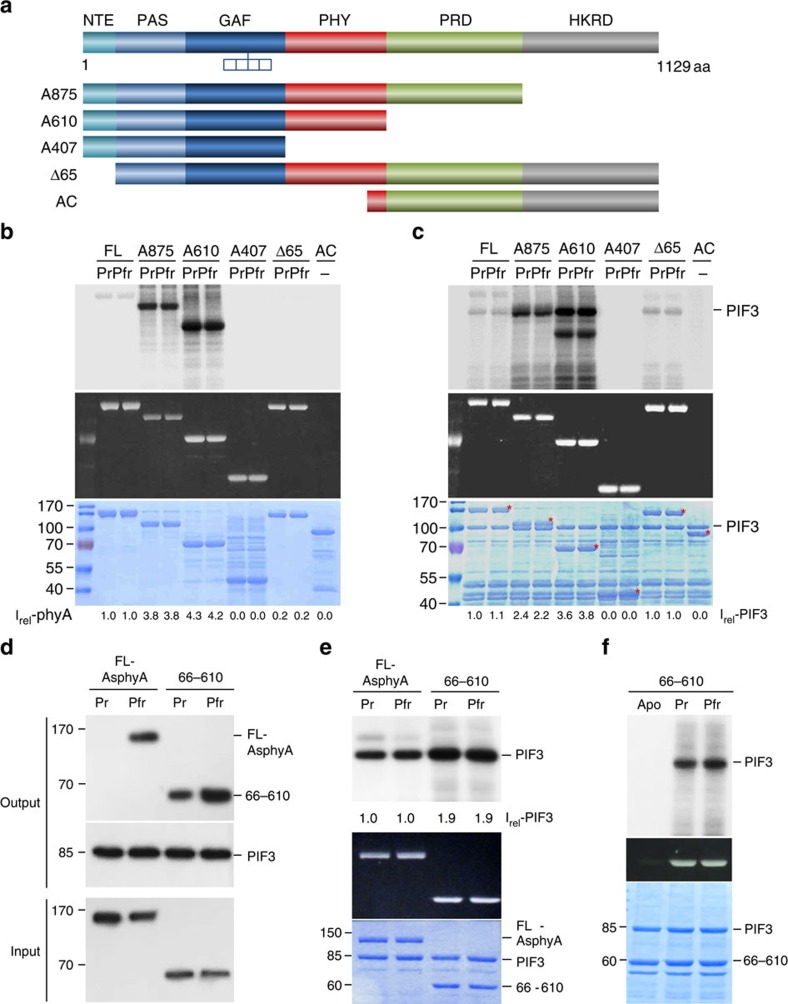
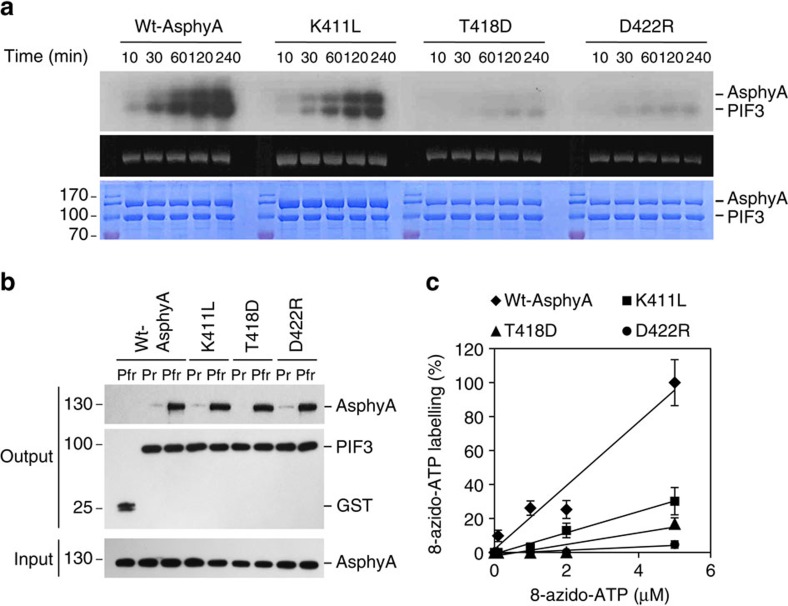
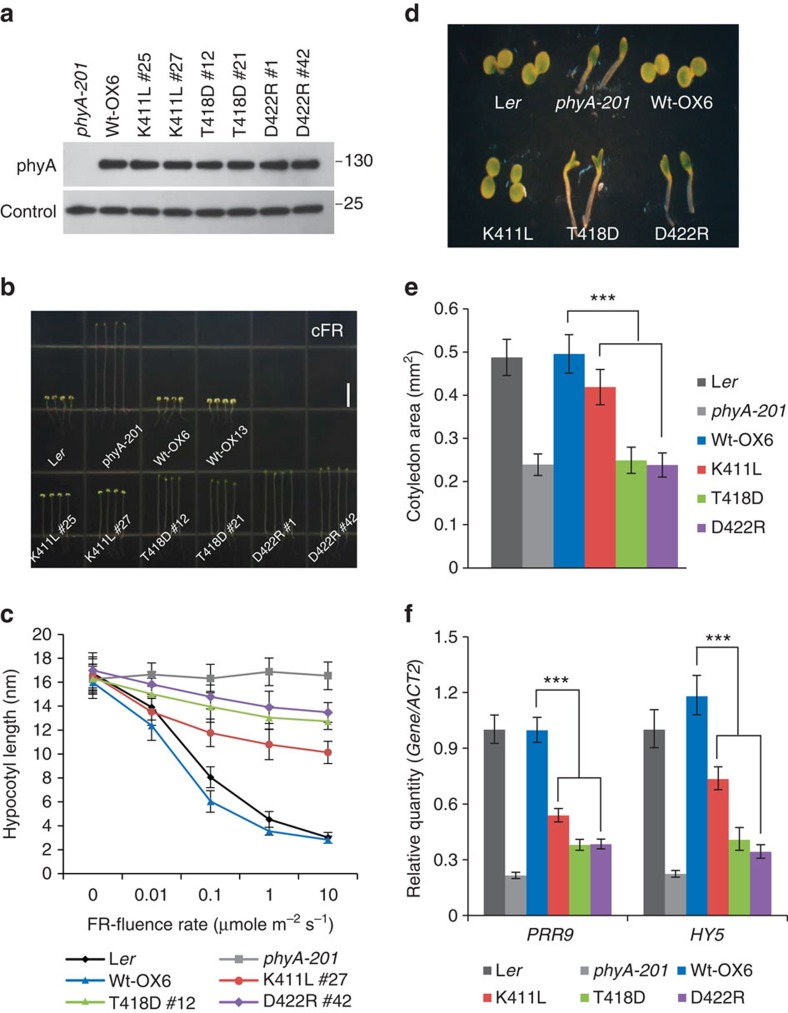
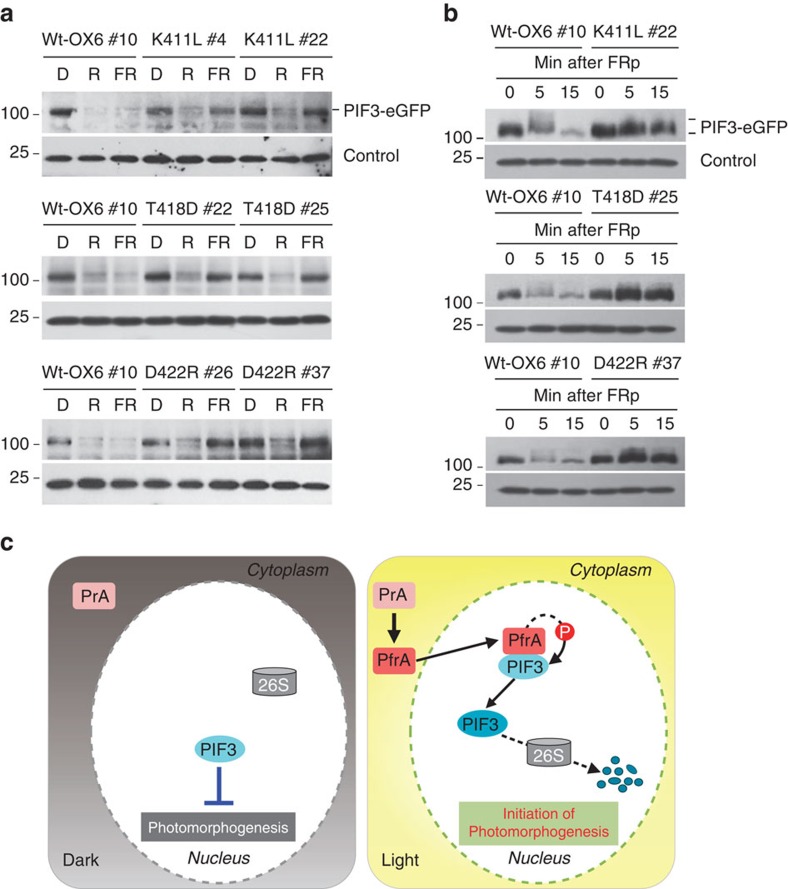
It has been suggested that plant phytochromes are autophosphorylating serine/threonine kinases. However, the biochemical properties and functional roles of putative phytochrome kinase activity in plant light signalling are largely unknown. Here, we describe the biochemical and functional characterization of Avena sativa phytochrome A (AsphyA) as a potential protein kinase. We provide evidence that phytochrome-interacting factors (PIFs) are phosphorylated by phytochromes in vitro. Domain mapping of AsphyA shows that the photosensory core region consisting of PAS-GAF-PHY domains in the N-terminal is required for the observed kinase activity. Moreover, we demonstrate that transgenic plants expressing mutant versions of AsphyA, which display reduced activity in in vitro kinase assays, show hyposensitive responses to far-red light. Further analysis reveals that far-red light-induced phosphorylation and degradation of PIF3 are significantly reduced in these transgenic plants. Collectively, these results suggest a positive relationship between phytochrome kinase activity and photoresponses in plants.
Figures
References
-
- Bae G. & Choi G. Decoding of light signals by plant phytochromes and their interacting proteins. Annu. Rev. Plant Biol. 59, 281–311 (2008). - PubMed
-
- Fankhauser C. & Chen M. Transposing phytochrome into the nucleus. Trends Plant Sci. 13, 596–601 (2008). - PubMed
-
- Henriques R., Jang I. C. & Chua N. H. Regulated proteolysis in light-related signaling pathways. Curr. Opin. Plant Biol. 12, 49–56 (2009). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
