

Domestication-driven Gossypium profilin 1 (GhPRF1) gene transduces early flowering phenotype in tobacco by spatial alteration of apical/floral-meristem related gene expression
- PMID: 27177585
- PMCID: PMC4866011
- DOI: 10.1186/s12870-016-0798-0
Domestication-driven Gossypium profilin 1 (GhPRF1) gene transduces early flowering phenotype in tobacco by spatial alteration of apical/floral-meristem related gene expression
Abstract
Background: Plant profilin genes encode core cell-wall structural proteins and are evidenced for their up-regulation under cotton domestication. Notwithstanding striking discoveries in the genetics of cell-wall organization in plants, little is explicit about the manner in which profilin-mediated molecular interplay and corresponding networks are altered, especially during cellular signalling of apical meristem determinacy and flower development.
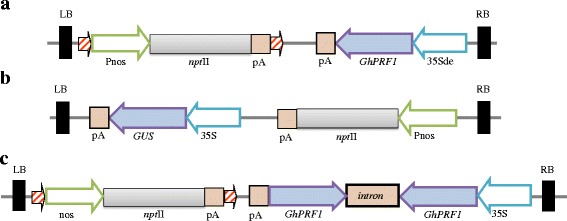
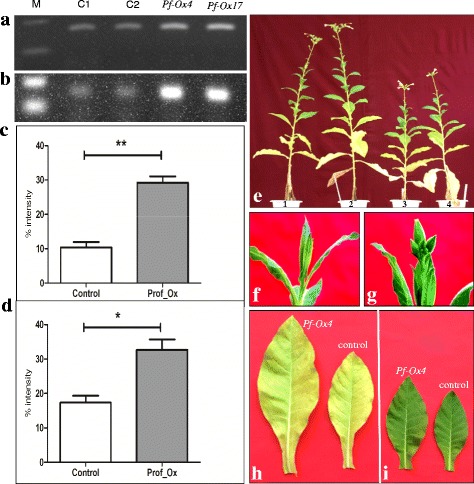
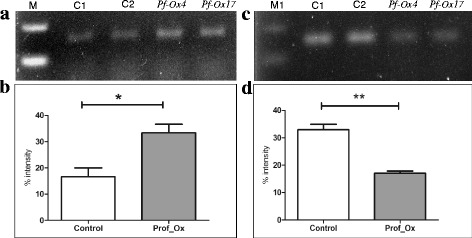
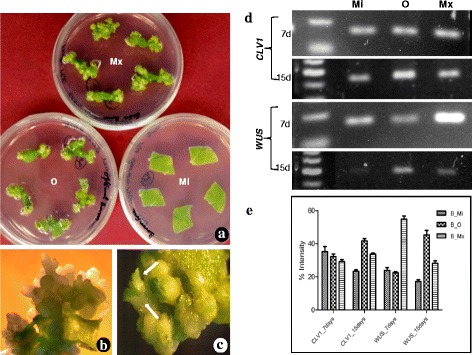
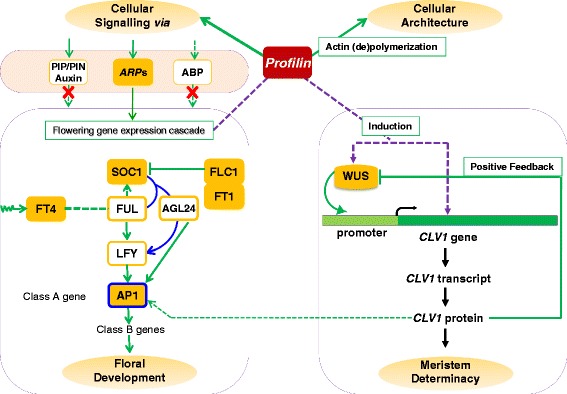
Results: Here we show that the ectopic expression of GhPRF1 gene in tobacco resulted in the hyperactivation of apical meristem and early flowering phenotype with increased flower number in comparison to the control plants. Spatial expression alteration in CLV1, a key meristem-determinacy gene, is induced by the GhPRF1 overexpression in a WUS-dependent manner and mediates cell signalling to promote flowering. But no such expression alterations are recorded in the GhPRF1-RNAi lines. The GhPRF1 transduces key positive flowering regulator AP1 gene via coordinated expression of FT4, SOC1, FLC1 and FT1 genes involved in the apical-to-floral meristem signalling cascade which is consistent with our in silico profilin interaction data. Remarkably, these positive and negative flowering regulators are spatially controlled by the Actin-Related Protein (ARP) genes, specifically ARP4 and ARP6 in proximate association with profilins. This study provides a novel and systematic link between GhPRF1 gene expression and the flower primordium initiation via up-regulation of the ARP genes, and an insight into the functional characterization of GhPRF1 gene acting upstream to the flowering mechanism. Also, the transgenic plants expressing GhPRF1 gene show an increase in the plant height, internode length, leaf size and plant vigor.
Conclusions: Overexpression of GhPRF1 gene induced early and increased flowering in tobacco with enhanced plant vigor. During apical meristem determinacy and flower development, the GhPRF1 gene directly influences key flowering regulators through ARP-genes, indicating for its role upstream in the apical-to-floral meristem signalling cascade.
Keywords: Apical meristem determinacy; Flower development; Flowering genes; Gene expression; Profilin.
Figures

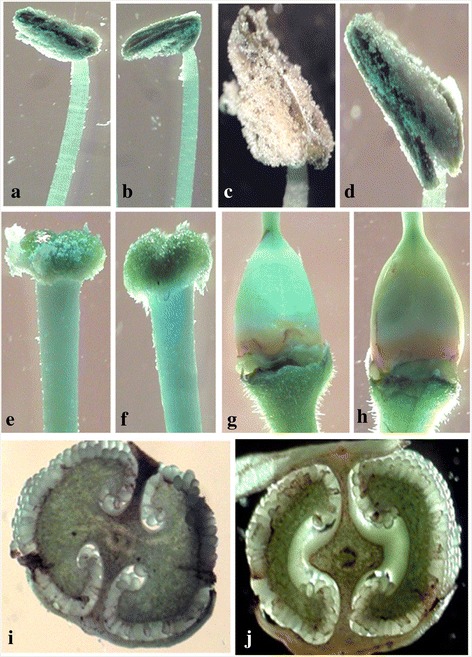
References
-
- Wan JM, Jiang L, Tang JY, Wang CM, Hou MY, Jing W, Zhang LX. Genetic dissection of the seed dormancy trait in cultivated rice (Oryza sativa L.) Plant Sci. 2005;170:786–92. doi: 10.1016/j.plantsci.2005.11.011. - DOI
-
- Blumler M. Modelling the origins of legume domestication and cultivation. Economic Bot. 1991;45:243–50. doi: 10.1007/BF02862051. - DOI
-
- Plitman U, Kislev M. Reproductive changes induced by domestication. In: Stirton C, Zarucchi J, editors. Advances in legume biology. St. Louis: Missouri Botanical Garden; 1989. pp. 487–503.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
