

RNA Duplex Map in Living Cells Reveals Higher-Order Transcriptome Structure
- PMID: 27180905
- PMCID: PMC5029792
- DOI: 10.1016/j.cell.2016.04.028
RNA Duplex Map in Living Cells Reveals Higher-Order Transcriptome Structure
Abstract
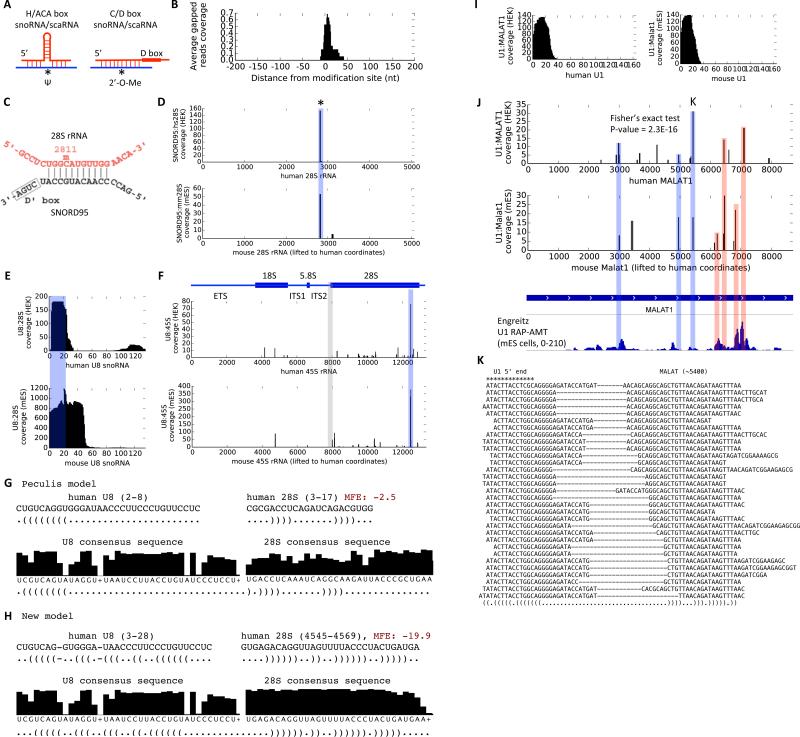
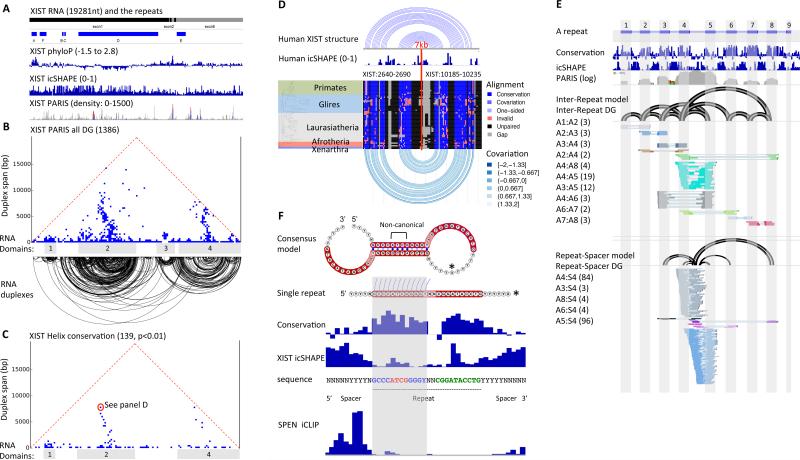
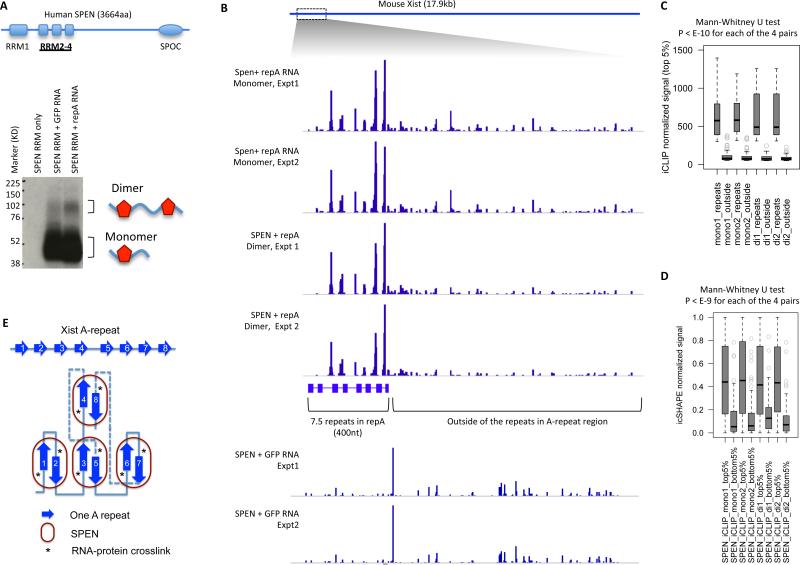
RNA has the intrinsic property to base pair, forming complex structures fundamental to its diverse functions. Here, we develop PARIS, a method based on reversible psoralen crosslinking for global mapping of RNA duplexes with near base-pair resolution in living cells. PARIS analysis in three human and mouse cell types reveals frequent long-range structures, higher-order architectures, and RNA-RNA interactions in trans across the transcriptome. PARIS determines base-pairing interactions on an individual-molecule level, revealing pervasive alternative conformations. We used PARIS-determined helices to guide phylogenetic analysis of RNA structures and discovered conserved long-range and alternative structures. XIST, a long noncoding RNA (lncRNA) essential for X chromosome inactivation, folds into evolutionarily conserved RNA structural domains that span many kilobases. XIST A-repeat forms complex inter-repeat duplexes that nucleate higher-order assembly of the key epigenetic silencing protein SPEN. PARIS is a generally applicable and versatile method that provides novel insights into the RNA structurome and interactome. VIDEO ABSTRACT.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
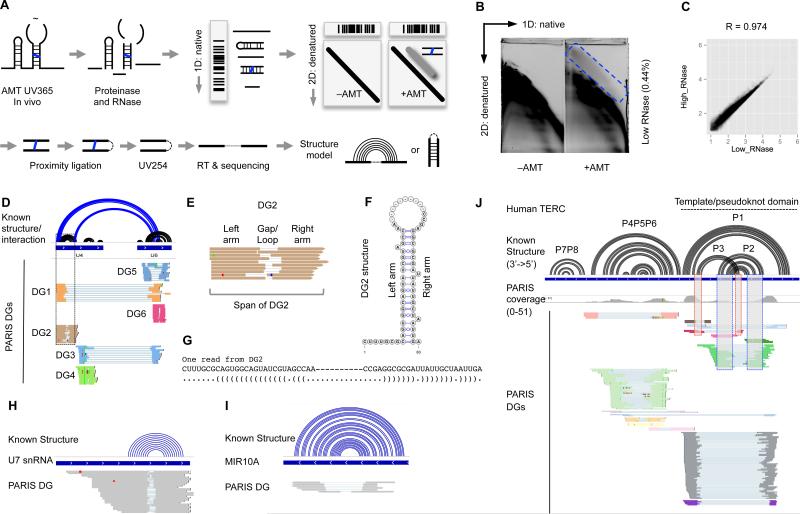
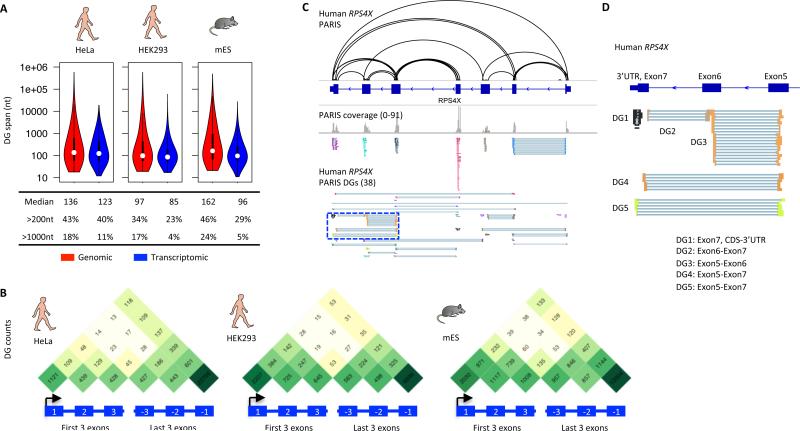
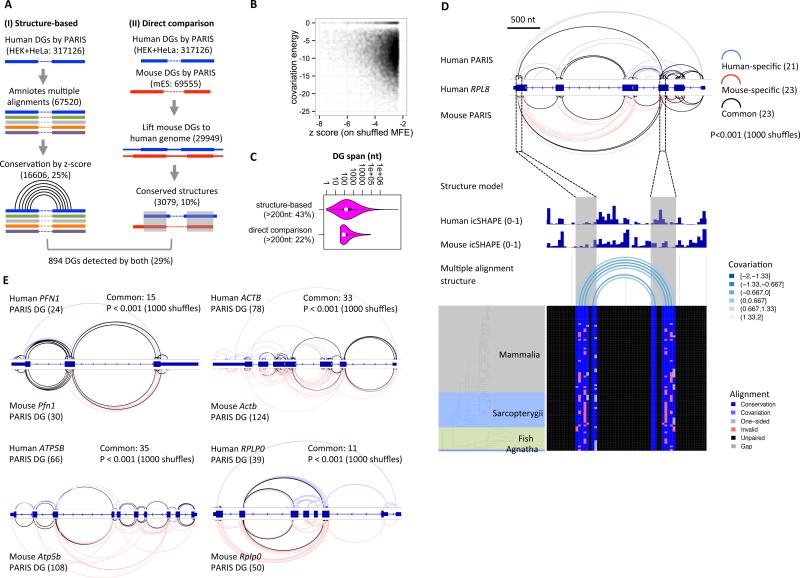
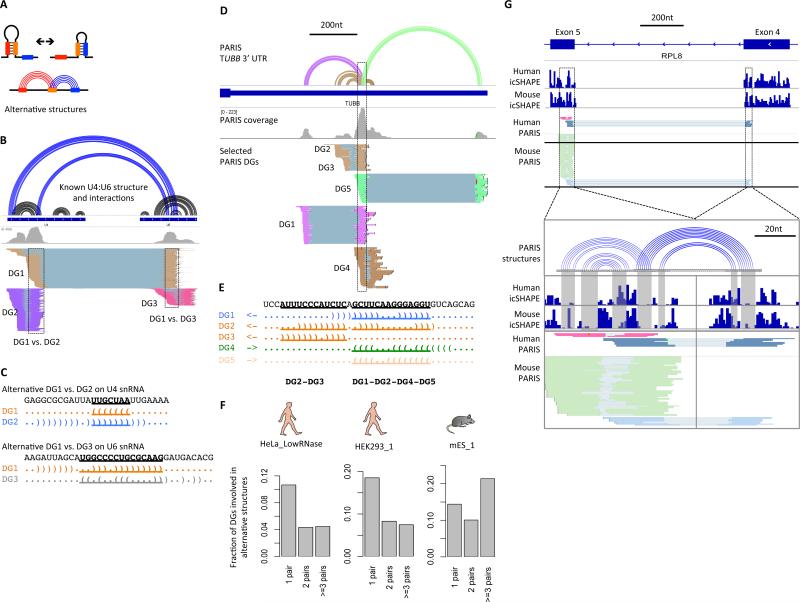
Comment in
-
Direct Duplex Detection: An Emerging Tool in the RNA Structure Analysis Toolbox.Trends Biochem Sci. 2016 Sep;41(9):734-736. doi: 10.1016/j.tibs.2016.07.001. Epub 2016 Jul 14. Trends Biochem Sci. 2016. PMID: 27427309 Free PMC article.
References
-
- Bejerano G, Pheasant M, Makunin I, Stephen S, Kent WJ, Mattick JS, Haussler D. Ultraconserved elements in the human genome. Science. 2004;304:1321–1325. - PubMed
-
- Brown CJ, Ballabio A, Rupert JL, Lafreniere RG, Grompe M, Tonlorenzi R, Willard HF. A gene from the region of the human X inactivation centre is expressed exclusively from the inactive X chromosome. Nature. 1991;349:38–44. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
