

Enzyme replacement with PEGylated cystathionine β-synthase ameliorates homocystinuria in murine model
- PMID: 27183385
- PMCID: PMC4887166
- DOI: 10.1172/JCI85396
Enzyme replacement with PEGylated cystathionine β-synthase ameliorates homocystinuria in murine model
Abstract
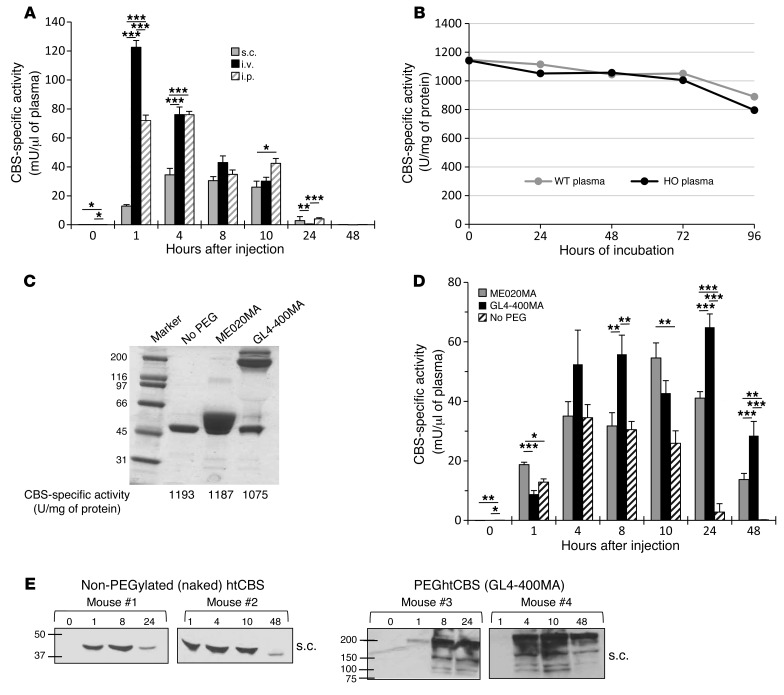
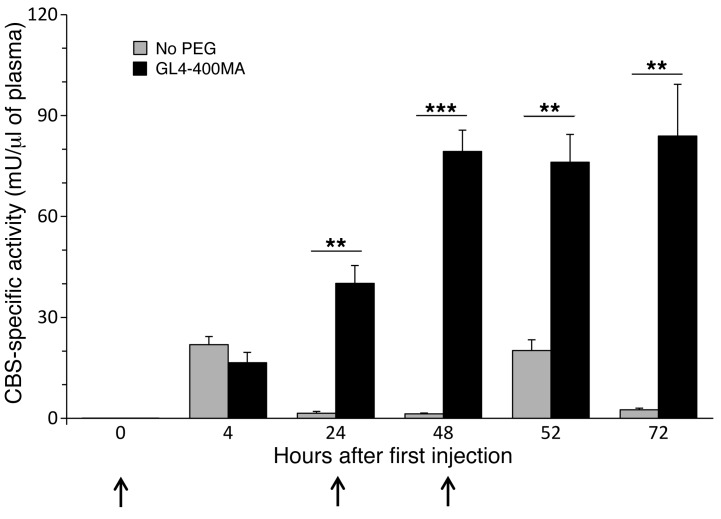
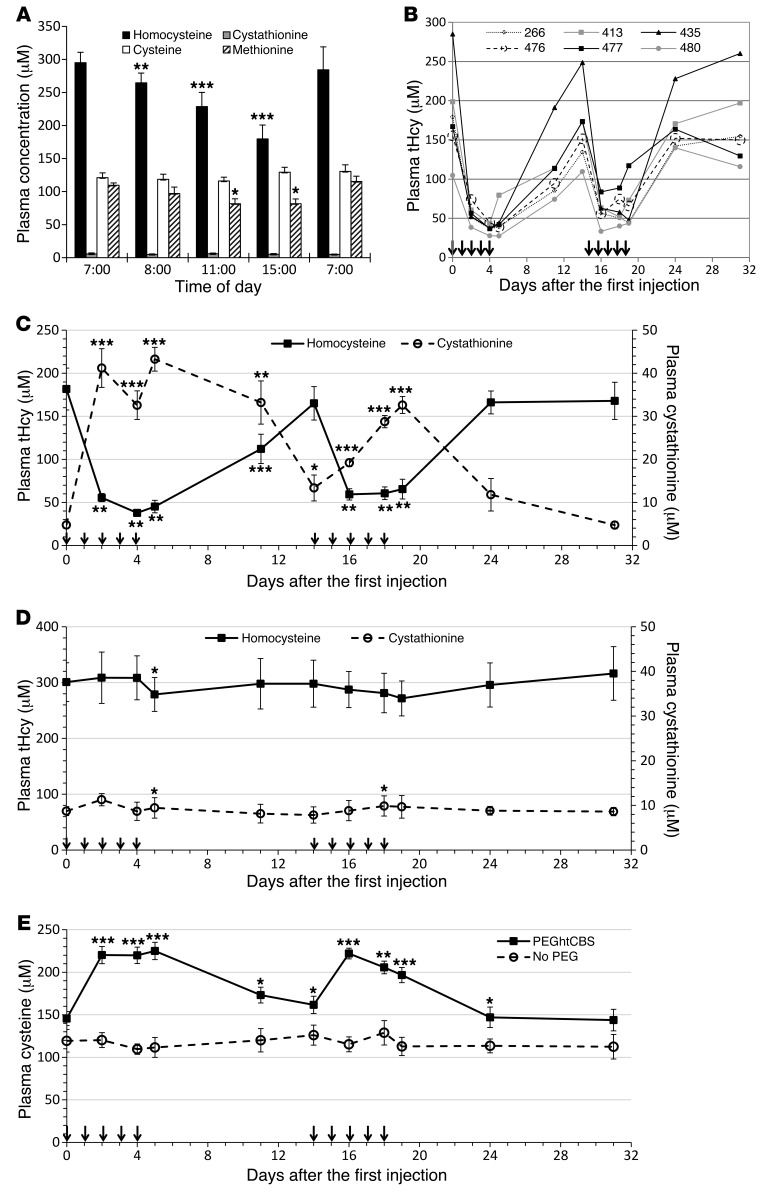
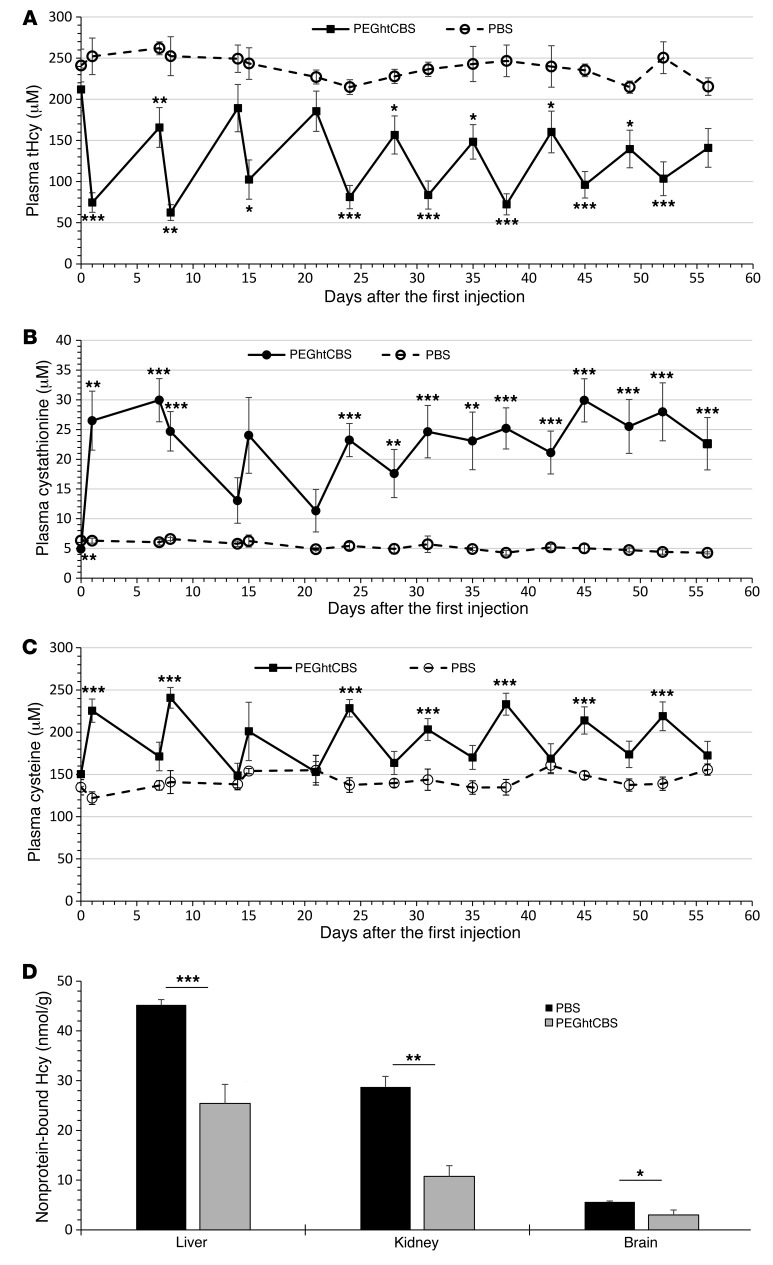
Homocystinuria, which typically results from cystathionine β-synthase (CBS) deficiency, is the most common defect of sulfur amino acid metabolism. CBS condenses homocysteine and serine to cystathionine that is then converted to cysteine. Individuals with homocystinuria have markedly elevated plasma levels of homocysteine and methionine and reduced concentrations of cystathionine and cysteine. Clinical disease manifestations include thromboembolism and neuropsychiatric, ocular, and skeletal complications. Here, we have shown that administration of PEGylated CBS into the circulation of homocystinuria model mice alters the extra- and intracellular equilibrium of sulfur amino acids, resulting in a decrease of approximately 75% in plasma total homocysteine (tHcy) and normalization of cysteine concentrations. Moreover, the decrease in homocysteine and the normalization of cysteine in PEGylated CBS-treated model mice were accompanied by improvement of histopathological liver symptoms and increased survival. Together, these data suggest that CBS enzyme replacement therapy (ERT) is a promising approach for the treatment of homocystinuria and that ERT for metabolic diseases may not necessitate introduction of the deficient enzyme into its natural intracellular compartment.
Figures
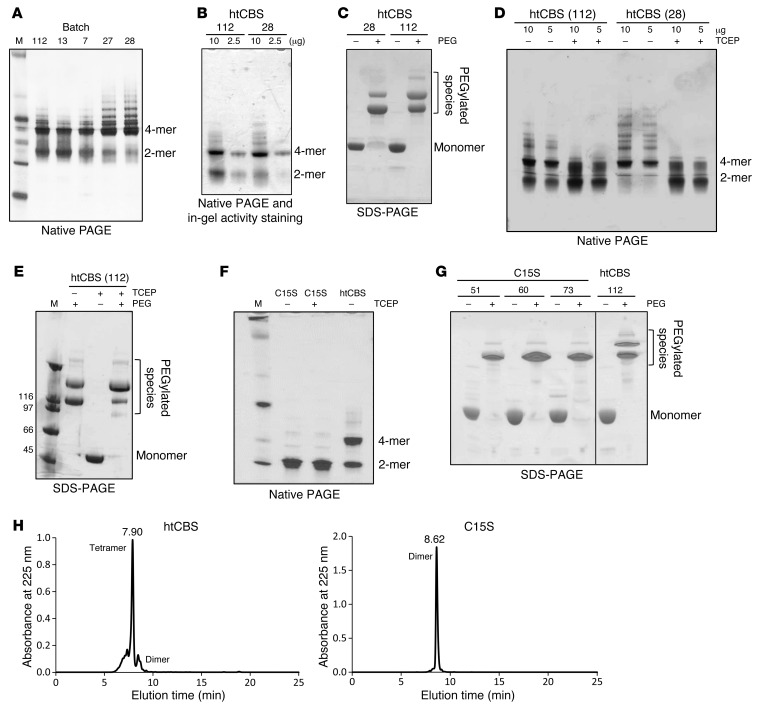
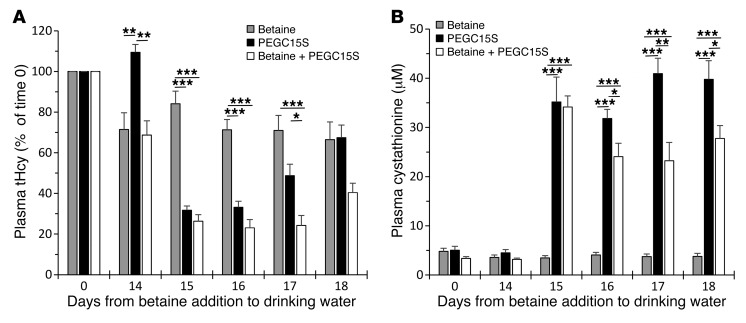
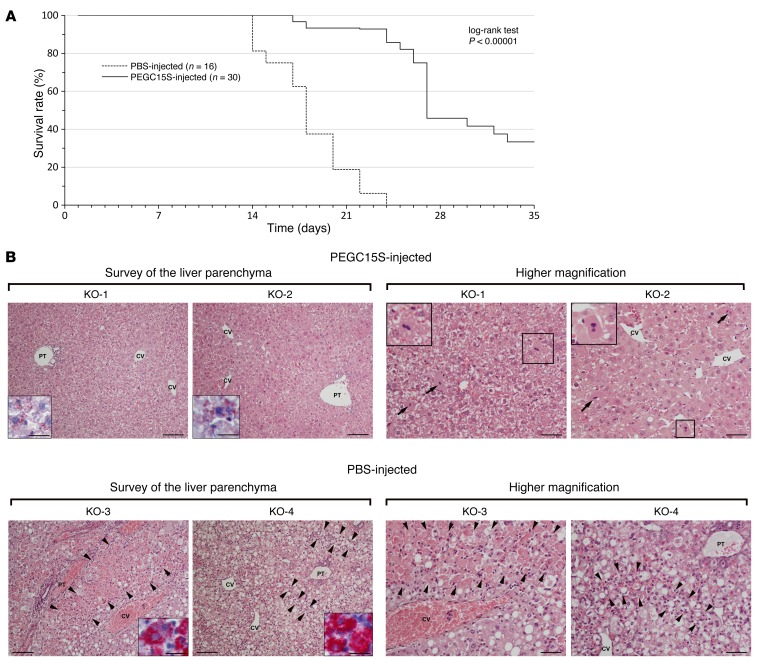
Comment in
-
Vision of correction for classic homocystinuria.J Clin Invest. 2016 Jun 1;126(6):2043-4. doi: 10.1172/JCI88251. Epub 2016 May 16. J Clin Invest. 2016. PMID: 27183384 Free PMC article.
References
-
- Mudd SH, Levy HL, Kraus JP. Disorders of transsulfuration. In: Scriver CR, et al., eds. The Metabolic and Molecular Bases of Inherited Disease. New York, New York, USA: McGraw-Hill; 2001:2007–2056.
-
- Kraus JP, Kožich V. Cystathionine-β-synthase and its deficiency. In: Carmel R, Jacobsen DW, editors. Homocysteine in Health and Disease. Cambridge, United Kingdom: Cambridge University Press; 2001. pp. 223–243. , eds.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
