

Haploinsufficiency of the 22q11.2 microdeletion gene Mrpl40 disrupts short-term synaptic plasticity and working memory through dysregulation of mitochondrial calcium
- PMID: 27184122
- PMCID: PMC5114177
- DOI: 10.1038/mp.2016.75
Haploinsufficiency of the 22q11.2 microdeletion gene Mrpl40 disrupts short-term synaptic plasticity and working memory through dysregulation of mitochondrial calcium
Abstract
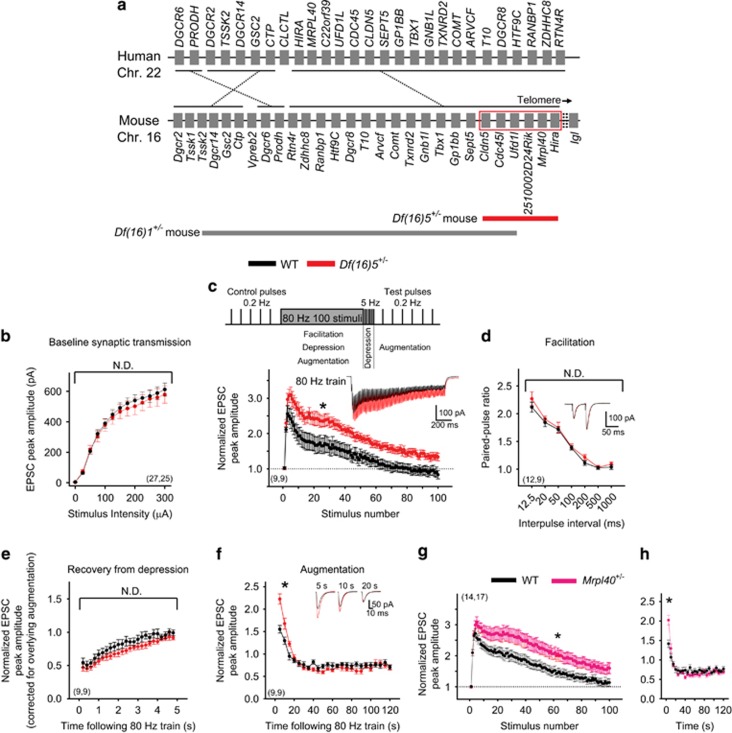
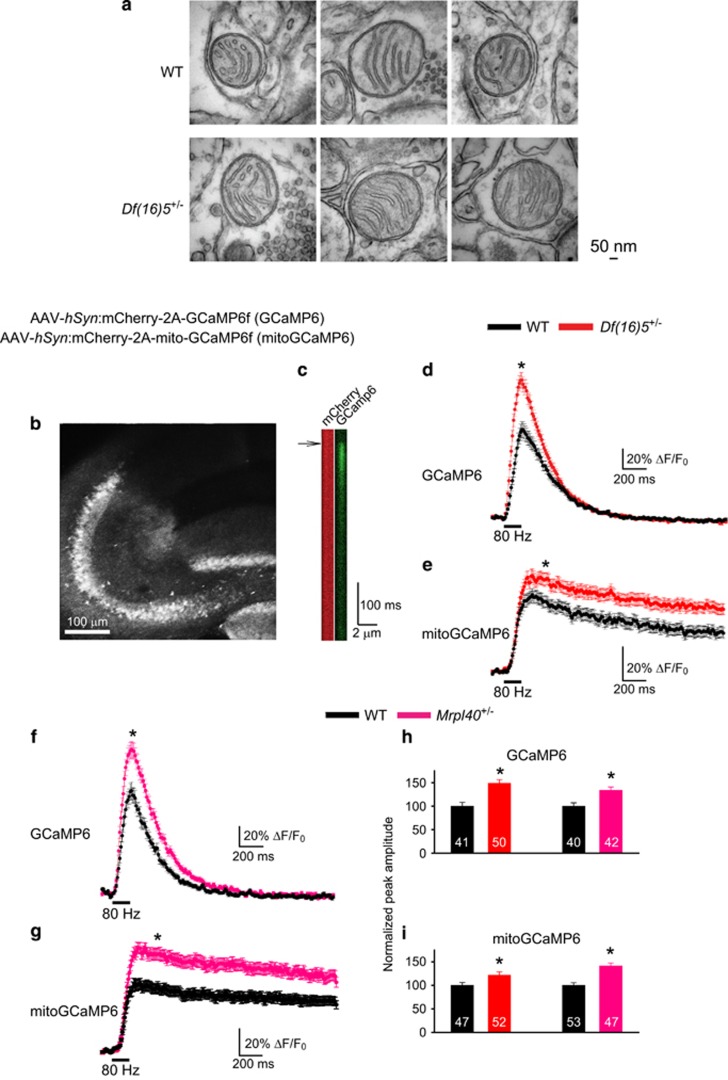
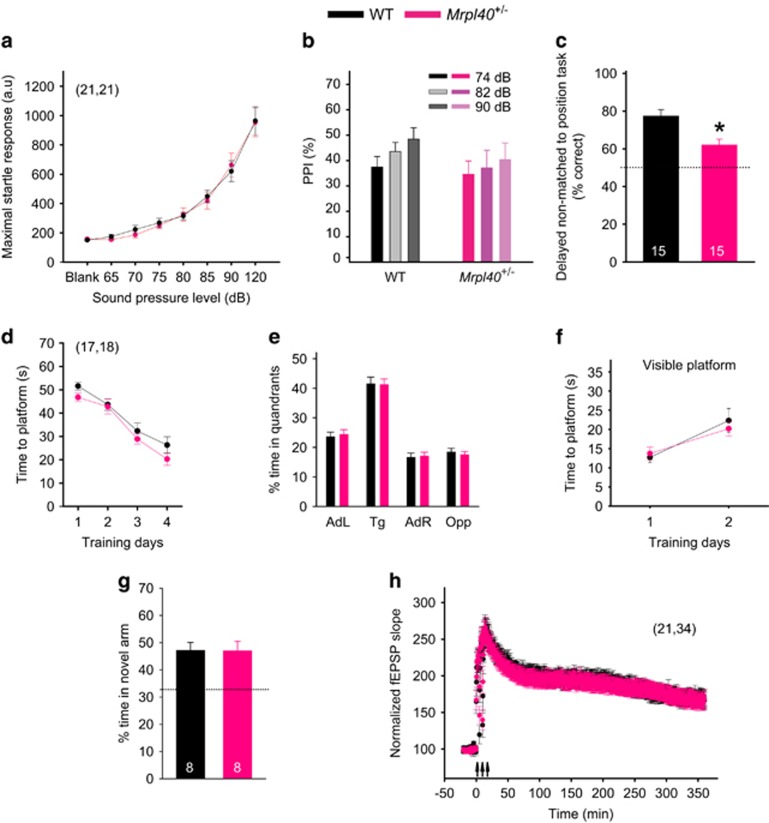
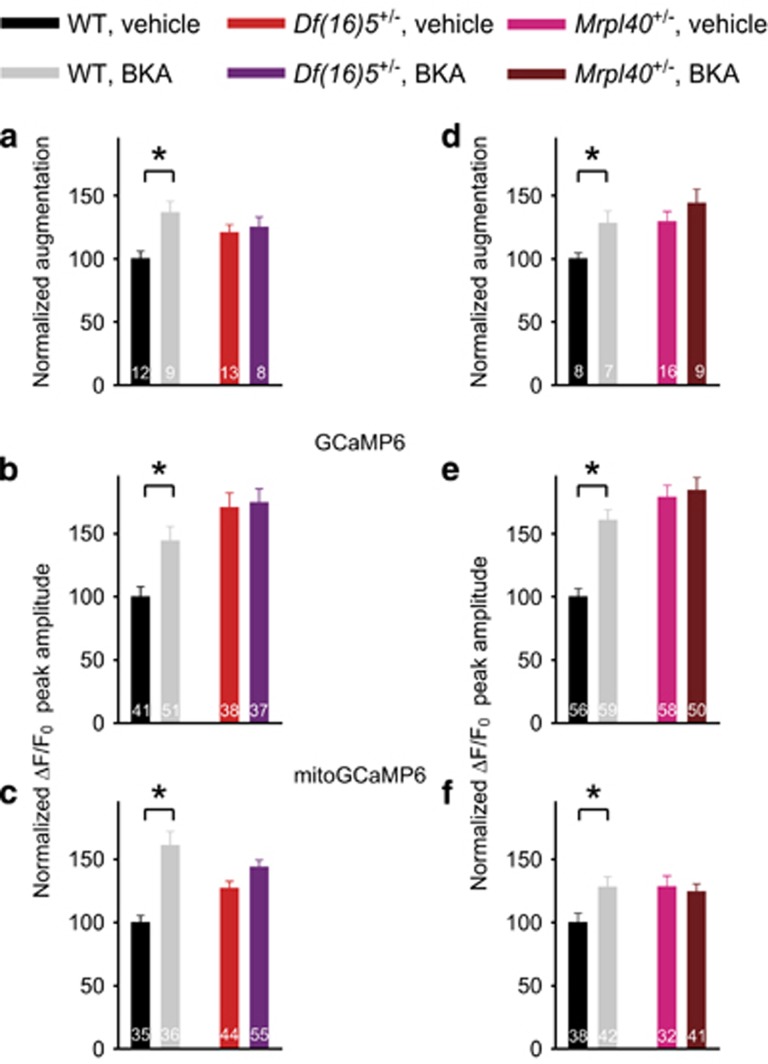
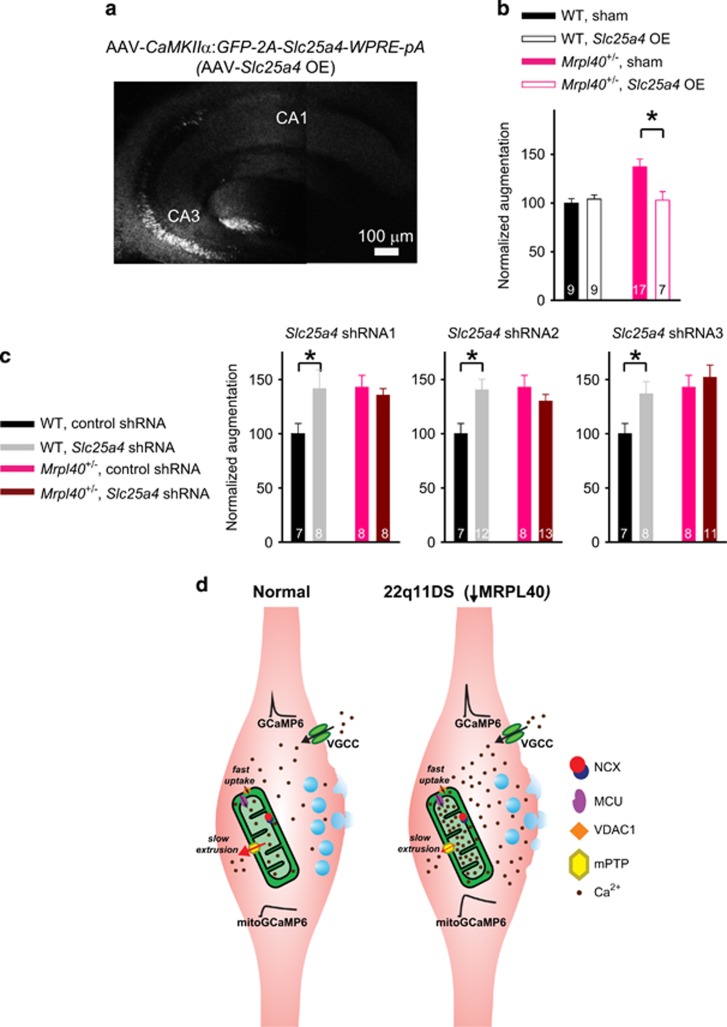
Hemizygous deletion of a 1.5- to 3-megabase region on chromosome 22 causes 22q11.2 deletion syndrome (22q11DS), which constitutes one of the strongest genetic risks for schizophrenia. Mouse models of 22q11DS have abnormal short-term synaptic plasticity that contributes to working-memory deficiencies similar to those in schizophrenia. We screened mutant mice carrying hemizygous deletions of 22q11DS genes and identified haploinsufficiency of Mrpl40 (mitochondrial large ribosomal subunit protein 40) as a contributor to abnormal short-term potentiation (STP), a major form of short-term synaptic plasticity. Two-photon imaging of the genetically encoded fluorescent calcium indicator GCaMP6, expressed in presynaptic cytosol or mitochondria, showed that Mrpl40 haploinsufficiency deregulates STP via impaired calcium extrusion from the mitochondrial matrix through the mitochondrial permeability transition pore. This led to abnormally high cytosolic calcium transients in presynaptic terminals and deficient working memory but did not affect long-term spatial memory. Thus, we propose that mitochondrial calcium deregulation is a novel pathogenic mechanism of cognitive deficiencies in schizophrenia.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Carpenter WT Jr, Buchanan RW. Schizophrenia. N Engl J Med 1994; 330: 681–690. - PubMed
-
- Scambler PJ. The 22q11 deletion syndromes. Hum Mol Genet 2000; 9: 2421–2426. - PubMed
-
- Pulver AE, Nestadt G, Goldberg R, Shprintzen RJ, Lamacz M, Wolyniec PS et al. Psychotic illness in patients diagnosed with velo-cardio-facial syndrome and their relatives. J Nerv Ment Dis 1994; 182: 476–478. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
