

A novel "complement-metabolism-inflammasome axis" as a key regulator of immune cell effector function
- PMID: 27184294
- PMCID: PMC5025719
- DOI: 10.1002/eji.201546131
A novel "complement-metabolism-inflammasome axis" as a key regulator of immune cell effector function
Abstract
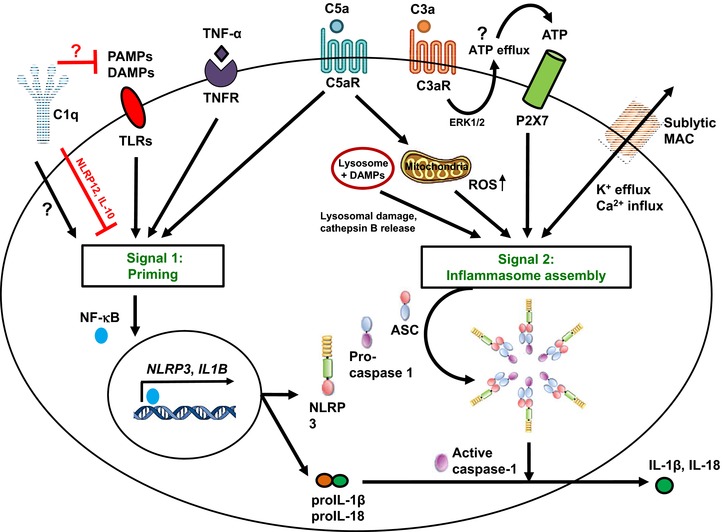
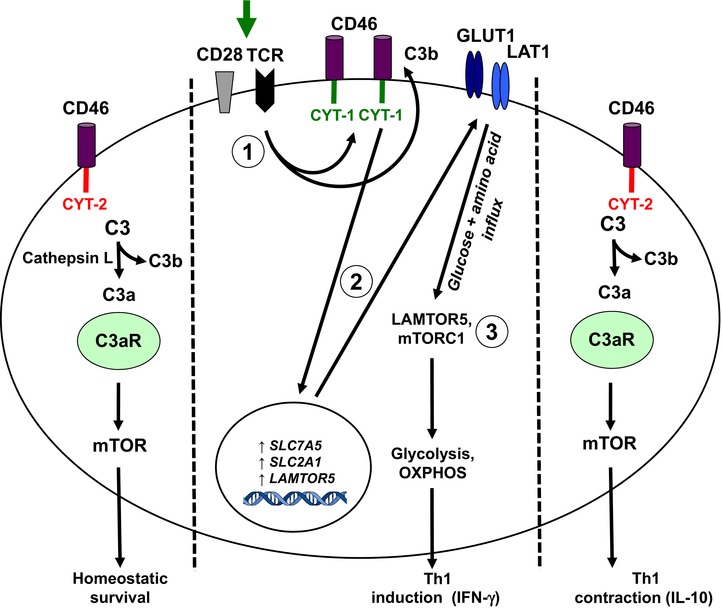
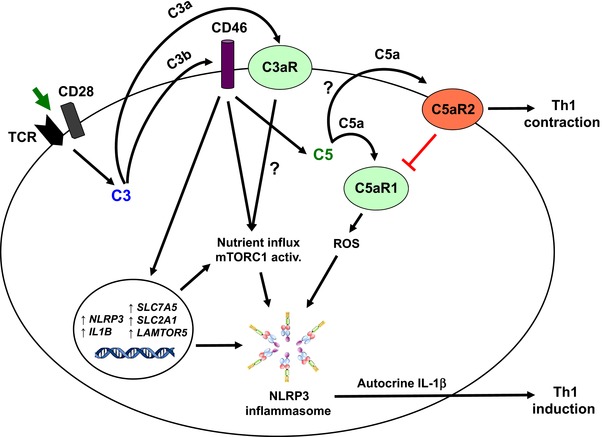
The inflammasomes are intracellular multiprotein complexes that induce and regulate the generation of the key pro-inflammatory cytokines IL-1β and IL-18 in response to infectious microbes and cellular stress. The activation of inflammasomes involves several upstream signals including classic pattern or danger recognition systems such as the TLRs. Recently, however, the activation of complement receptors, such as the anaphylatoxin C3a and C5a receptors and the complement regulator CD46, in conjunction with the sensing of cell metabolic changes, for instance increased amino acid influx and glycolysis (via mTORC1), have emerged as additional critical activators of the inflammasome. This review summarizes recent advances in our knowledge about complement-mediated inflammasome activation, with a specific focus on a novel "complement - metabolism - NLRP3 inflammasome axis."
Keywords: Complement; Metabolism; NLRP3 inflammasome.
© 2016 The Authors. European Journal of Immunology published by WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Figures
References
-
- Creagh, E. M. and O'Neill, L. A. , TLRs, NLRs and RLRs: a trinity of pathogen sensors that co‐operate in innate immunity. Trends Immunol. 2006. 8: 352–357. - PubMed
-
- Köhl, J. , The role of complement in danger sensing and transmission. Immunol. Res. 2006. 34: 157–176. - PubMed
-
- Dowling, J. K. and O'Neill, L. A. , Biochemical regulation of the inflammasome. Crit. Rev. Biochem. Mol. Biol. 2012. 47: 424–443. - PubMed
-
- Asgari, E. , Le Friec, G. , Yamamoto, H. , Perucha, E. , Sacks, S. S. , Köhl, J. , Cook, H. T. and Kemper, C. , C3a modulates IL‐1β secretion in human monocytes by regulating ATP efflux and subsequent NLRP3 inflammasome activation. Blood 2013. 122: 3473–3481. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
