

Regulation of Irregular Neuronal Firing by Autaptic Transmission
- PMID: 27185280
- PMCID: PMC4869121
- DOI: 10.1038/srep26096
Regulation of Irregular Neuronal Firing by Autaptic Transmission
Abstract
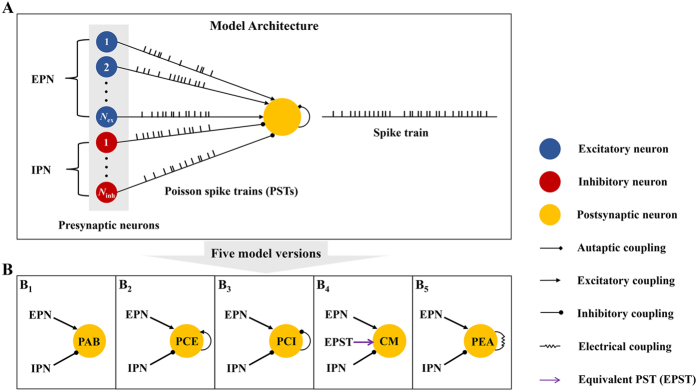
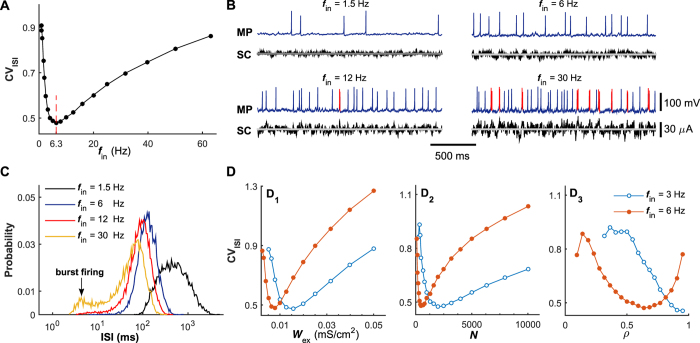
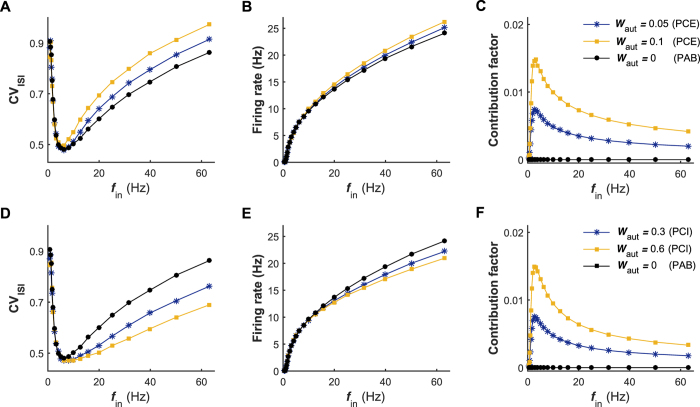
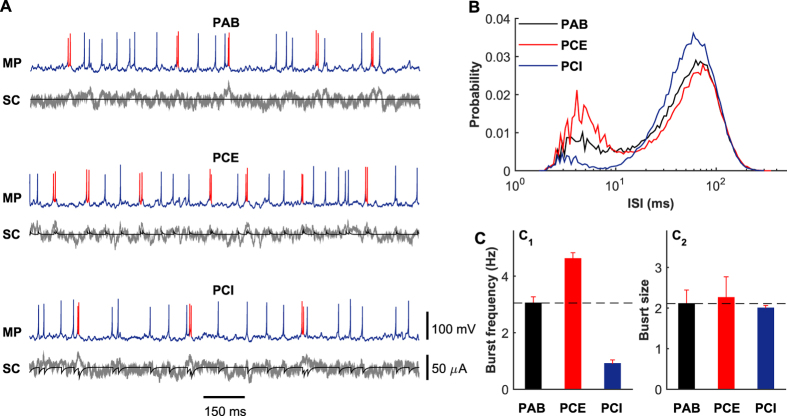
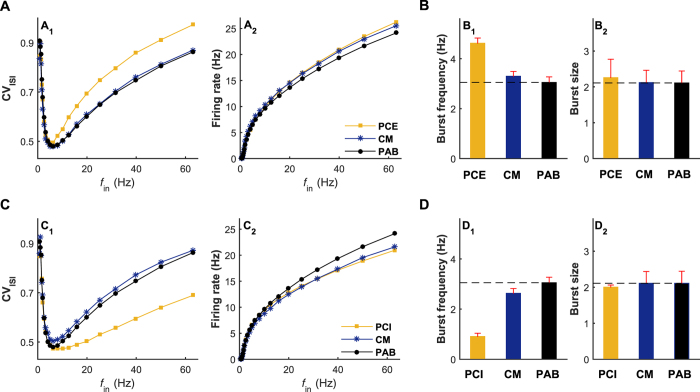
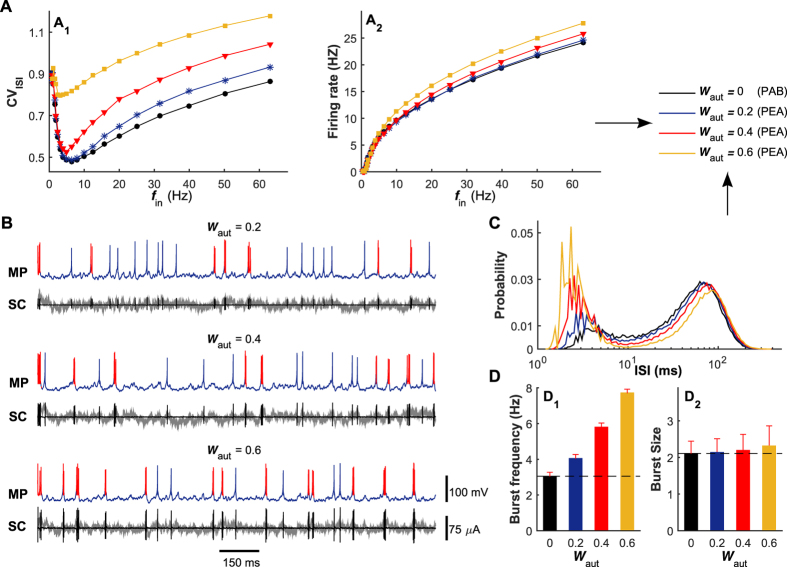
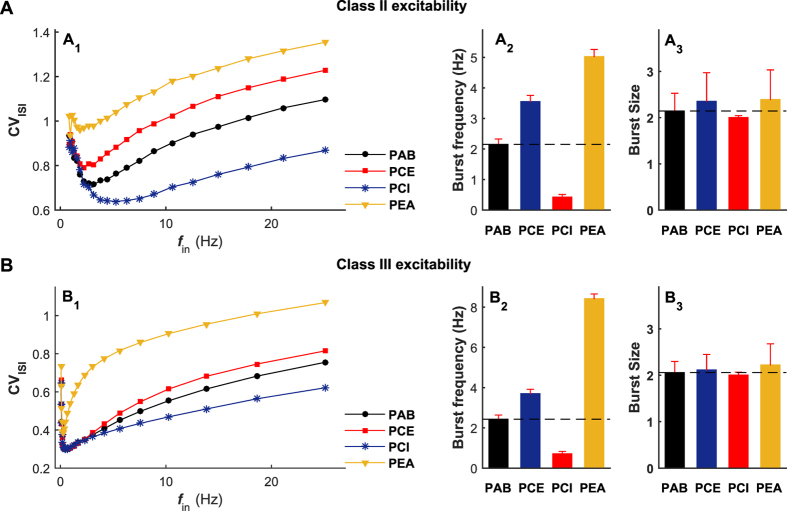
The importance of self-feedback autaptic transmission in modulating spike-time irregularity is still poorly understood. By using a biophysical model that incorporates autaptic coupling, we here show that self-innervation of neurons participates in the modulation of irregular neuronal firing, primarily by regulating the occurrence frequency of burst firing. In particular, we find that both excitatory and electrical autapses increase the occurrence of burst firing, thus reducing neuronal firing regularity. In contrast, inhibitory autapses suppress burst firing and therefore tend to improve the regularity of neuronal firing. Importantly, we show that these findings are independent of the firing properties of individual neurons, and as such can be observed for neurons operating in different modes. Our results provide an insightful mechanistic understanding of how different types of autapses shape irregular firing at the single-neuron level, and they highlight the functional importance of autaptic self-innervation in taming and modulating neurodynamics.
Figures

Similar articles
-
Autaptic Connections Shift Network Excitability and Bursting.Sci Rep. 2017 Mar 7;7:44006. doi: 10.1038/srep44006. Sci Rep. 2017. PMID: 28266594 Free PMC article.
-
Control of bursting behavior in neurons by autaptic modulation.Neurol Sci. 2013 Nov;34(11):1977-84. doi: 10.1007/s10072-013-1429-2. Epub 2013 Apr 18. Neurol Sci. 2013. PMID: 23595543
-
Contribution of AMPA and NMDA receptors in the spontaneous firing patterns of single neurons in autaptic culture.Biosystems. 2020 Dec;198:104278. doi: 10.1016/j.biosystems.2020.104278. Epub 2020 Oct 17. Biosystems. 2020. PMID: 33075473
-
Biophysical models of intrinsic homeostasis: Firing rates and beyond.Curr Opin Neurobiol. 2021 Oct;70:81-88. doi: 10.1016/j.conb.2021.07.011. Epub 2021 Aug 25. Curr Opin Neurobiol. 2021. PMID: 34454303 Review.
-
Neuronal firing: does function follow form?Curr Biol. 1996 Dec 1;6(12):1560-2. doi: 10.1016/s0960-9822(02)70771-9. Curr Biol. 1996. PMID: 8994812 Review.
Cited by
-
Stimulus-induced Epileptic Spike-Wave Discharges in Thalamocortical Model with Disinhibition.Sci Rep. 2016 Nov 23;6:37703. doi: 10.1038/srep37703. Sci Rep. 2016. PMID: 27876879 Free PMC article.
-
Effects of synaptic integration on the dynamics and computational performance of spiking neural network.Cogn Neurodyn. 2020 Jun;14(3):347-357. doi: 10.1007/s11571-020-09572-y. Epub 2020 Feb 19. Cogn Neurodyn. 2020. PMID: 32399076 Free PMC article.
-
Stimulus-induced transitions between spike-wave discharges and spindles with the modulation of thalamic reticular nucleus.J Comput Neurosci. 2017 Dec;43(3):203-225. doi: 10.1007/s10827-017-0658-4. Epub 2017 Sep 22. J Comput Neurosci. 2017. PMID: 28939929
-
Morphology controls how hippocampal CA1 pyramidal neuron responds to uniform electric fields: a biophysical modeling study.Sci Rep. 2017 Jun 12;7(1):3210. doi: 10.1038/s41598-017-03547-6. Sci Rep. 2017. PMID: 28607422 Free PMC article.
-
Dendritic and Axonal Propagation Delays Determine Emergent Structures of Neuronal Networks with Plastic Synapses.Sci Rep. 2017 Jan 3;7:39682. doi: 10.1038/srep39682. Sci Rep. 2017. PMID: 28045109 Free PMC article.
References
-
- Destexhe A. & Rudolph-Lilith M. Neuronal Noise (Springer, 2012).
-
- Balenzuela P. & Garca-Ojalvo J. Role of chemical synapses in coupled neurons with noise. Phys. Rev. E 72, 021901 (2005). - PubMed
-
- Zaikin A., Garca-Ojalvo J., Báscones R., Ullner E. & Kurths J. Doubly stochastic coherence via noise-induced symmetry in bistable neural models. Phys. Rev. Lett. 90(3), 030601 (2003). - PubMed
-
- Zhou C. & Kurths J. Noise-induced synchronization and coherence resonance of a Hodgkin-Huxley model of thermally sensitive neurons. Chaos 13(1), 401–409 (2003). - PubMed
-
- Stevens C. F. & Zador A. M. Input synchrony and the irregular firing of cortical neurons. Nat. Neurosci. 1(3), 210–217 (1998). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
