

CEP295 interacts with microtubules and is required for centriole elongation
- PMID: 27185865
- PMCID: PMC4958302
- DOI: 10.1242/jcs.186338
CEP295 interacts with microtubules and is required for centriole elongation
Abstract
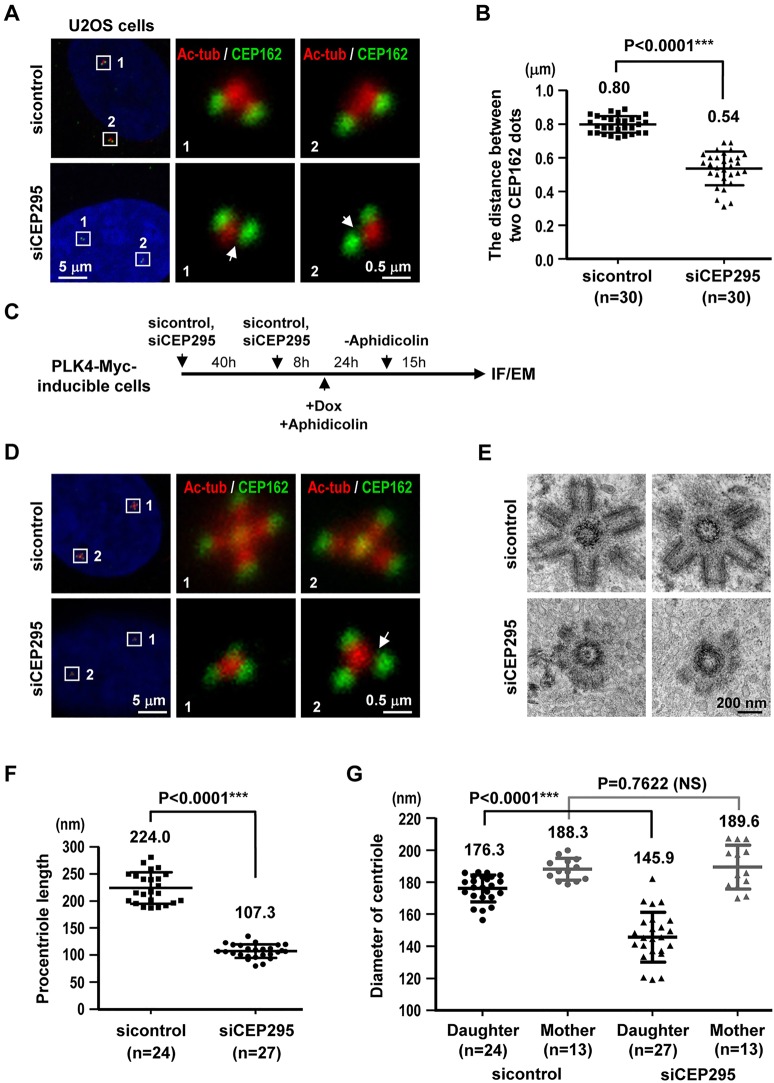
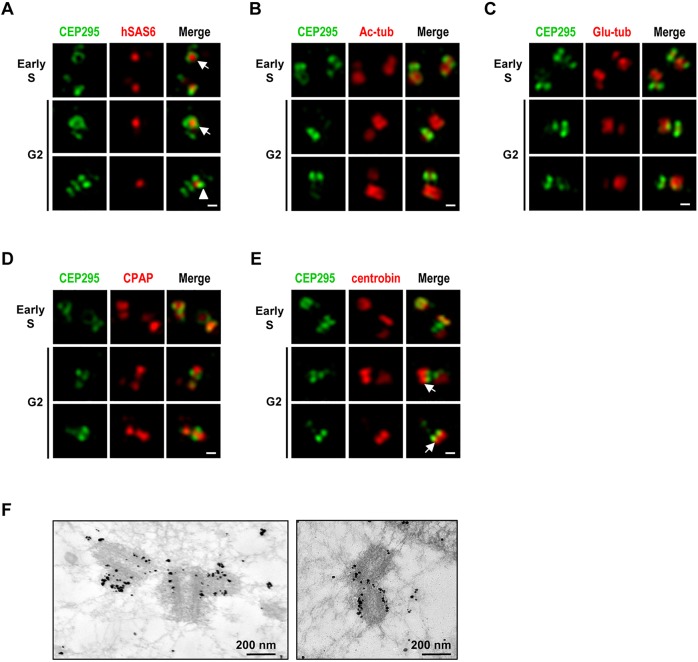
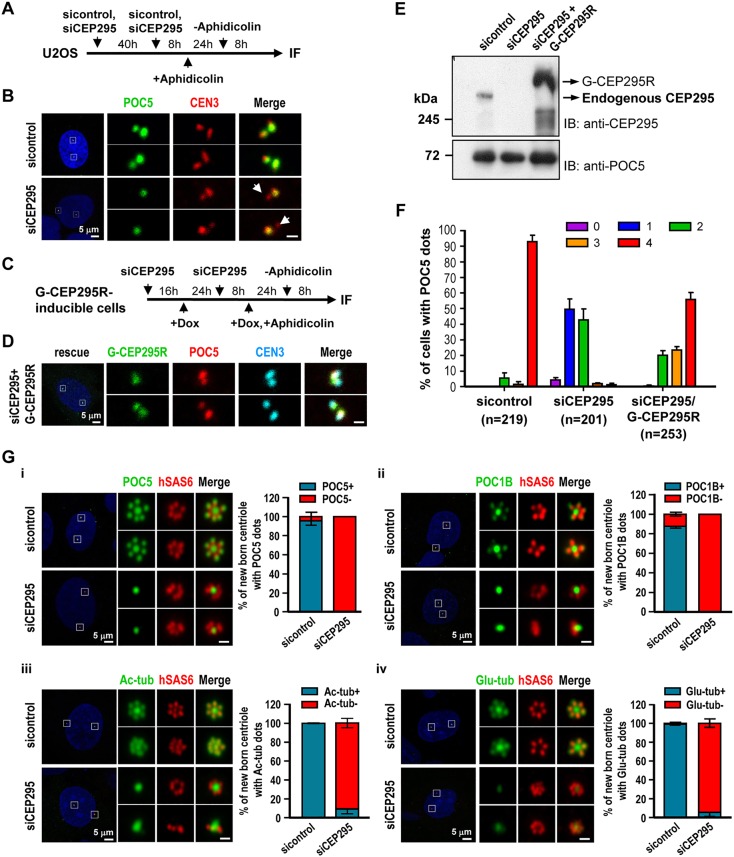
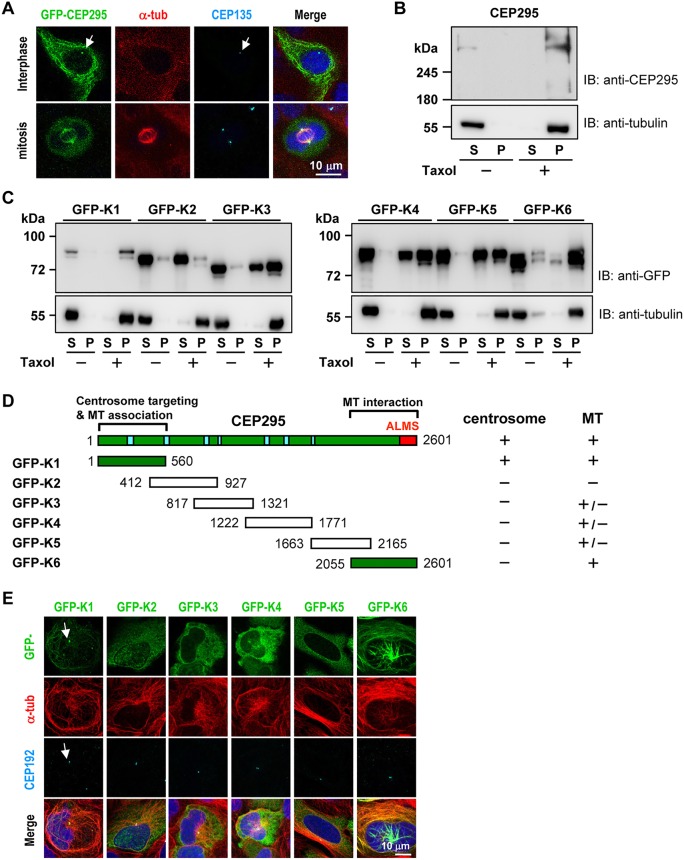
Centriole duplication is a tightly ordered process during which procentrioles are assembled in G1-S and elongate during S and G2. Here, we show that human CEP295 (Drosophila Ana1) is not essential for initial cartwheel assembly, but is required to build distal half centrioles during S and G2. Using super-resolution and immunogold electron microscopy, we demonstrate that CEP295 is recruited to the proximal end of procentrioles in early S phase, when it is also localized at the centriolar microtubule wall that surrounds the human SAS6 cartwheel hub. Interestingly, depletion of CEP295 not only inhibits the recruitments of POC5 and POC1B to the distal half centrioles in G2, resulting in shorter centrioles, it also blocks the post-translational modification of centriolar microtubules (e.g. acetylation and glutamylation). Importantly, our results indicate that CEP295 directly interacts with microtubules, and that excess CEP295 could induce the assembly of overly long centrioles. Furthermore, exogenous expression of the N-terminal domain of CEP295 exerts a dominant-negative effect on centriole elongation. Collectively, these findings suggest that CEP295 is essential for building the distal half centrioles and for post-translational modification of centriolar microtubules.
Keywords: Acetylation; Centriole assembly; Centriole duplication; Centrosome; Polyglutamylation; Procentriole formation.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
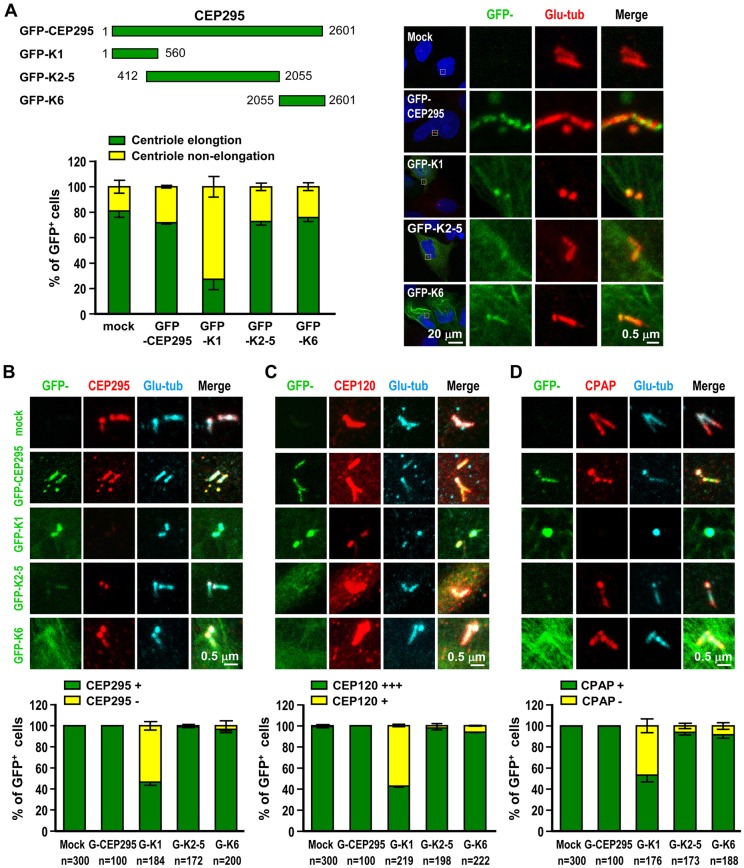
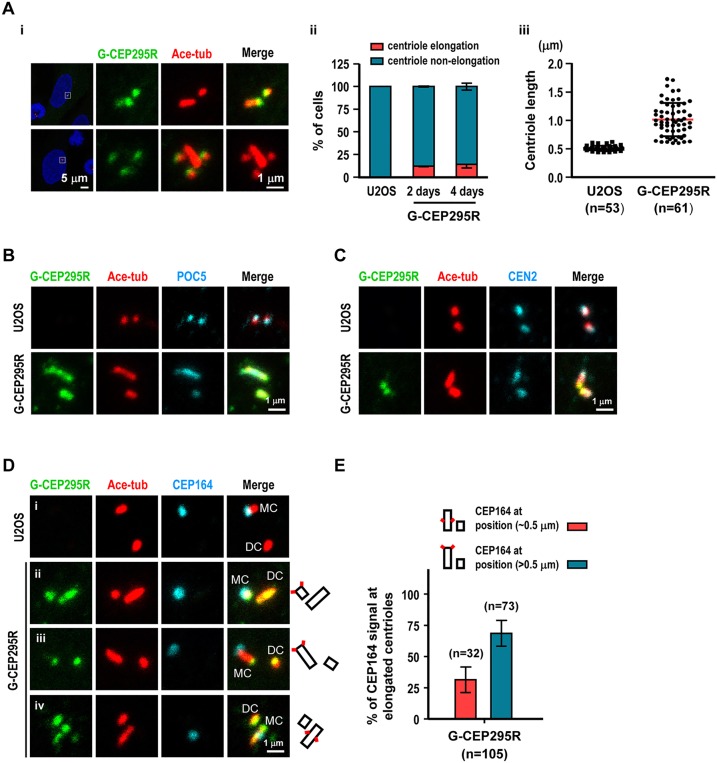
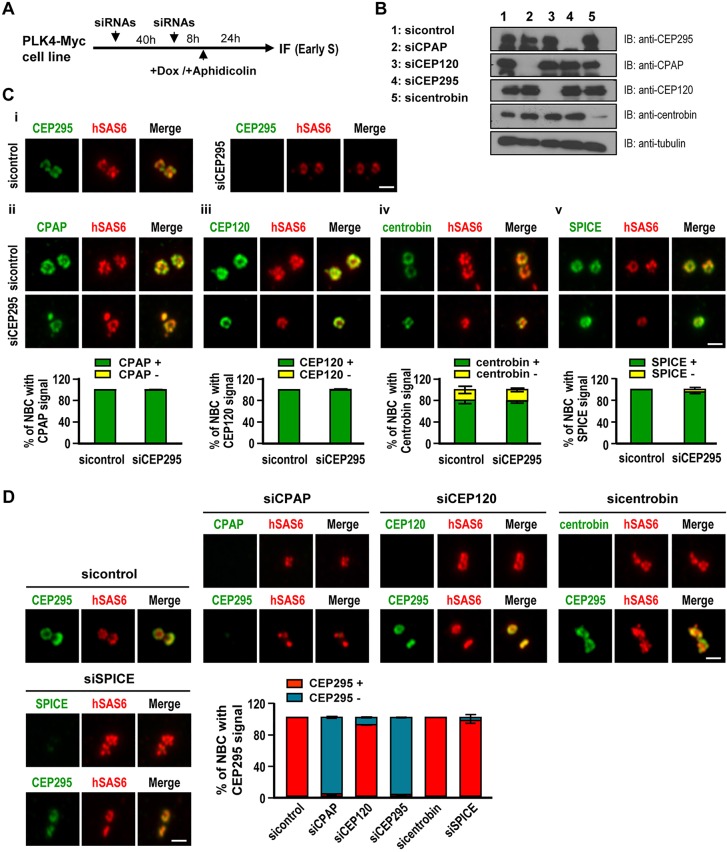
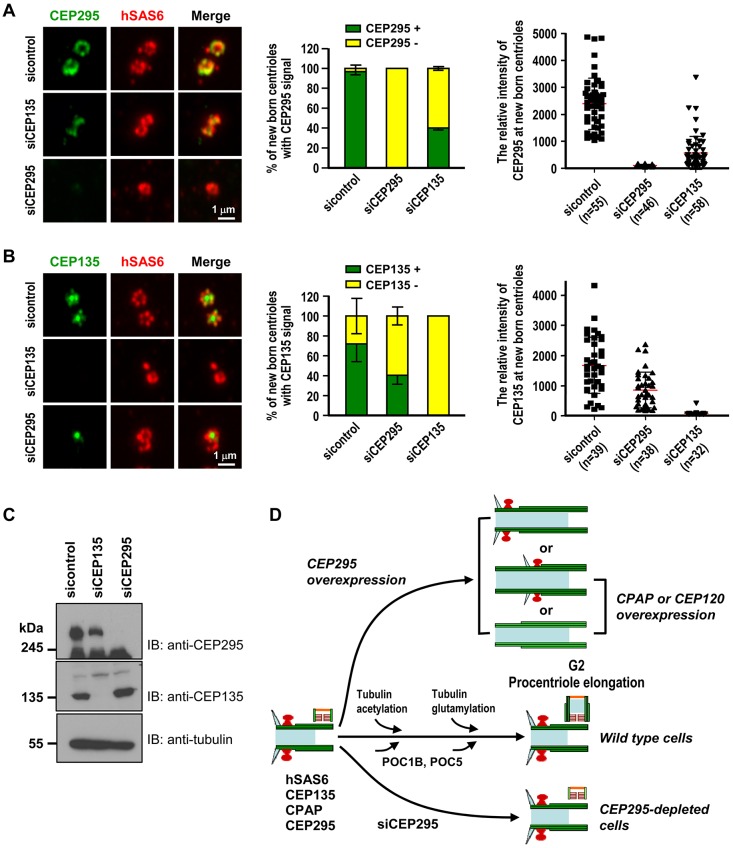
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
